DE60114830T2 - Verfahren zur herstellung modifizierter glycoproteine - Google Patents
Verfahren zur herstellung modifizierter glycoproteine Download PDFInfo
- Publication number
- DE60114830T2 DE60114830T2 DE60114830T DE60114830T DE60114830T2 DE 60114830 T2 DE60114830 T2 DE 60114830T2 DE 60114830 T DE60114830 T DE 60114830T DE 60114830 T DE60114830 T DE 60114830T DE 60114830 T2 DE60114830 T2 DE 60114830T2
- Authority
- DE
- Germany
- Prior art keywords
- host cell
- golgi
- glcnac
- glycoprotein
- glycosylation
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Revoked
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/005—Glycopeptides, glycoproteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/08—Drugs for disorders of the metabolism for glucose homeostasis
- A61P3/10—Drugs for disorders of the metabolism for glucose homeostasis for hyperglycaemia, e.g. antidiabetics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
- C12N15/81—Vectors or expression systems specially adapted for eukaryotic hosts for fungi for yeasts
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2477—Hemicellulases not provided in a preceding group
- C12N9/2488—Mannanases
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
- C12Y302/01113—Mannosyl-oligosaccharide 1,2-alpha-mannosidase (3.2.1.113), i.e. alpha-1,2-mannosidase
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/01—Fusion polypeptide containing a localisation/targetting motif
- C07K2319/04—Fusion polypeptide containing a localisation/targetting motif containing an ER retention signal such as a C-terminal HDEL motif
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/01—Fusion polypeptide containing a localisation/targetting motif
- C07K2319/05—Fusion polypeptide containing a localisation/targetting motif containing a GOLGI retention signal
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biomedical Technology (AREA)
- Microbiology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Mycology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- Diabetes (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Virology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Botany (AREA)
- Endocrinology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Pain & Pain Management (AREA)
- Rheumatology (AREA)
- Hematology (AREA)
- Obesity (AREA)
- Immunology (AREA)
- Emergency Medicine (AREA)
Description
- HINTERGRUND DER ERFINDUNG
- Glycosylierungswege
- Neu synthetisierte Proteine können in Zellen einer weiteren Prozessierung unterliegen, die als posttranslationale Modifikaton bekannt ist. Im Besonderen können Zuckerreste enzymatisch hinzugefügt werden, ein Prozess, der als Glycosylierung bekannt ist. Die resultierenden Proteine, die kovalent gekoppelte Oligosaccharid-Seitenketten tragen, sind als glycosylierte Proteine oder Glycoproteine bekannt. Bakterien glycosylieren Proteine im Allgemeinen nicht; in Zellen, in denen Glycosylierung vorkommt, tritt sie im Allgemeinen an unspezifischen Stellen in dem Protein auf (Moens and Vanderleyden, Arch. Microbiol. 1997 168 (3): 169–175).
- Eukaryonten hängen im Allgemeinen ein spezifisches Oligosaccharid an die Seitenkette eines Protein-Asparaginrests an, im Besonderen an ein Asparagin, das in der Sequenz Asn-Xaa-Ser/Thr/Cys vorkommt (wobei Xaa eine beliebige Aminosäure darstellt). Nach der Anheftung der Saccharid-Einheit, die als ein N-Glycan bekannt ist, können in vivo weitere Modifikationen auftreten. Im Allgemeinen treten diese Modifikationen in einer geordneten Folge enzymatischer Reaktionen auf, die als eine Kaskade bekannt sind. Verschiedene Organismen stellen unterschiedliche Glycosylierungs-Enzyme (Glycosyltransferasen und Glycosidasen) und verschiedene Glycosyl-Substrate bereit, sodass die endgültige Zusammensetzung einer Zucker-Seitenkette in Abhängigkeit von dem Wirt ausgesprochen stark variieren kann.
- Mikroorganismen wie filamentöse Pilze und Hefen (niedere Eukaryonten), hängen zum Beispiel im Allgemeinen zusätzliche Mannose- und/oder Mannosylphosphat-Zucker an. Das resultierende Glycan ist als "High-Mannose"-Typ oder als ein Mannan bekannt. Im Gegensatz dazu kann die im Entstehen begriffene Oligosaccharid-Seitenkette in tierischen Zellen zurechtgetrimmt werden, wobei einige Mannosereste entfernt werden und die Kette mit zusätzlichen Zuckerresten verlängert wird, die im Allgemeinen in den N-Glycanen von niederen Eukaryonten nicht auftreten. Siehe R. K. Bretthauer, et al. Biotechnology and Applied Biochemistry,; 1999, 30, 193–200; W. Martinet, et al. Biotechnology Letters, 1998, 20, 1171–1177; S. Weikert, et al. Nature Biotechnology Letters, 1999, 17, 1116–1121; M. Malissard, et al. Biochemical and Biophysical Research Communications, 2000, 267, 169–173; Jarvis, et al. 1998 Engineering N-glycosylation pathways in the baculovirus-insect cell system, Current Opinion in Biotechnology, 9: 528–533; und M. Takeuchi, 1997 Trends in Glycoscience and Glycotechnology, 1997, 9, S29–S35.
- Die N-Glycane, die in Menschen und Tieren hergestellt werden, werden im Allgemeinen als komplexe N-Glycane bezeichnet. Ein komplexes N-Glycan bedeutet eine Struktur mit im Allgemeinen zwei bis sechs äußeren Verzweigungen, wobei eine Sialyl-lactosamin-Sequenz an die innere Kernstruktur Man3GlcNAc2 gekoppelt ist. Ein komplexes N-Glycan hat mindestens eine Verzweigung, und vorzugsweise mindestens zwei, von sich abwechselnden GlcNAc- und Galactose (Gal)-Resten, die in Oligosacchariden enden, wie zum Beispiel: NeuNAc-; NeuAcα2-6GalNAcα1-; NeuAcα2-3Ga1β1-3GalNAcα1-; NeuAcα2-3/6Galβ1-4GlcNAcβ1-; GlcNAcα1-4Ga1β1-(ausschließlich Mucine); Fucα1-2Ga1β1-(Blutgruppe H). An Galactose-, GalNAc- und GlcNAc-Resten können Sulfatester auftreten und an Mannoseresten können Phosphatester auftreten. NeuAc (Neu: Neuraminsäure; Ac: Acetylrest) kann O-acetyliert oder durch NeuGl (N-Glycolylneuraminsäure) ausgetauscht sein. Komplexe N-Glycane können auch innerhalb der Kette Substitutionen aus zweiteilenden GlcNAc und Kern-Fucose (Fuc) tragen.
- Glycosylierung beim Menschen beginnt mit einem aufeinanderfolgenden Satz von Reaktionen im endoplasmatischen Retikulum (ER), was zu einer Kern-Oligosaccharid-Struktur führt, die an den Asparaginrest in der Sequenz Asn-Xaa-Ser/Thr von neu synthetisierten Proteinen übertragen wird (siehe
1A ). Im ER tritt eine weitere Prozessierung durch Glucosidasen und Mannosidasen auf, bevor das im Entstehen begriffene Glycoprotein in den frühen Golgi-Apparat übertragen wird, wo zusätzliche Mannosereste durch Golgi-spezifische 1,2-Mannosidasen entfernt werden. Die Prozessierung wird fortgesetzt, während das Protein durch den Golgi voranschreitet. In dem mittleren Golgi fügt eine Vielzahl modifizierender Enzyme, beinhaltend N-Acetylglucosamin-Transferasen (GnTI, GnTII, GnTIII, GnTIV, GnTV, GnTVI), Mannosidase II, Fucosyltransferasen, spezifische Zuckerreste and und entfernt diese (siehe1B ). Schließlich wirken in dem Trans-Golgi Galactosyltransferasen und Sialyltransferasen (ST) auf die N-Glycane und das fertiggestellte Glycoprotein wird aus dem Golgi-Apparat freigesetzt. Die Protein-N-Glycane von tierischen Glycoproteinen haben bi-, tri-, oder tetra-antennäre Strukturen und können im Allgemeinen Galactose, Fucose und N-Acetylglucosamin beinhalten. Im Allgemeinen bestehen die terminalen Reste der N-Glycane aus Sialinsäure. Eine typische Struktur eines menschlichen N-Glycans ist in1B gezeigt. - Zucker-Nucleotid-Vorläufer
- Die N-Glycane tierischer Glycoproteine beinhalten im Allgemeinen Galactose, Fucose und terminale Sialinsäure. Diese Zucker werden im Allgemeinen nicht an Glycoproteinen gefunden, die in Hefe und filamentösen Pilzen hergestellt werden. In Menschen wird im Allgemeinen der gesamte Bereich an Nucleotid-Zucker-Vorläufer (z.B. UDP-N-Acetylglucosamin, UDP-N-Acetylgalactosamin, CMP-N-Acetylneuraminsäure, UDP-Galactose, GDP-Fucose, etc.) im Cytosol synthetisiert und in den Golgi transportiert, wo sie durch Glycosyltransferasen an das Kern-Oligosaccharid angeheftet werden (Sommers and Hirschberg, 1981, J. Cell Biol. 91 (2): A409–A406; Sommers and Hirschberg 1982 J. Biol. Chem. 257 (18): 811–817; Perez and Hirschberg 1987 Methods in Enzymology 138: 709–715).
- Glycosyl-Transfer-Reaktionen bringen in der Regel ein Nebenprodukt hervor, bei dem es sich um ein Nucleosid-Diphosphat oder -Monophosphat handelt. Während Monophosphate im Austausch gegen Nucleosid-Triphosphat-Zucker durch einen Antiport-Mechanismus direkt exportiert werden können, müssen Diphospho-Nucleoside (z.B. GDP) durch Phosphatasen (z.B. GDPase) gespalten werden, wobei Nucleosid-Monophosphate und anorganisches Phosphat entstehen, bevor diese exportiert werden. Diese Reaktion ist für eine wirkungsvolle Glycosylierung wichtig; es zeigte sich zum Beispiel, dass GDPase aus S. cerevisiae für die Mannosylierung notwendig ist. Die GDPase hat jedoch gegenüber UDP eine um 90% reduzierte Aktivität (Berninsone et al., 1994 J. Biol. Chem. 269 (1): 207–211α). Niederen Eukaryonten fehlt im Allgemeinen UDP-spezifische Diphosphatase-Aktivität im Golgi, da diese für die Golgi-basierte Glycoprotein-Synthese keine UDP-Zucker-Vorläufer verwenden. Man fand heraus, dass Schizosaccharomyces pombe, eine Hefe, von der sich zeigte, dass sie Galactose-Reste an Zellwand-Polysaccharide (aus UDP-Galactose) anfügt, spezifische UDPase-Aktivität besitzt, was die Notwendigkeit eines solchen Enzyms anzeigt (Berninsone et al., 1994). UDP ist als ein starker Hemmstoff von Glycosyltransferasen bekannt und das Entfernen dieses Glycosylierungs-Nebenprodukts ist wichtig, um in dem Lumen des Golgi Glycosyltransferase-Hemmung zu verhindern (Khatara et al., 1974). Siehe Berninsone, P., et al., 1995 J. Biol. Chem. 270 (24): 14564–14567; Beaudet, L. et al. 1998 Abc Transporters: Biochemical, Cellular, and Molecular Aspects. 191: 397–413.
- Kompartimentierung von Glycosylierungs-Enzymen
- Glycosyltransferasen und Mannosidasen kleiden die innere (luminale) Oberfläche des ER und des Golgi-Apparates aus und stellen dabei eine katalytische Oberfläche bereit, die die sequenzielle Prozessierung von Glycoproteinen erlaubt, während diese durch das ER und das Golgi-Netzwerk fortschreiten. Die zahlreichen Kompartimente des Cis-, des mittleren und des Trans-Golgi und das Trans-Golgi-Netzwerk (TGN) stellen die verschiedenen Lokalitäten bereit, in denen die geordnete Abfolge von Glycosylierungs-Reaktionen stattfinden kann. Sobald ein Glycoprotein von der Synthese im ER bis hin zu seiner vollen Reife im späten Golgi oder TGN fortschreitet, wird es sequenziell verschiedenen Glycosidasen, Mannosidasen und Glycosyltransferasen ausgesetzt, sodass eine spezifische N-Glycan-Struktur synthetisiert werden kann. Die Enzyme beinhalten im Allgemeinen eine katalytische Domäne, eine Stammregion, eine Membran-durchdringende Region und einen N-terminalen cytoplasmatischen Schwanz. Die letzteren drei strukturellen Komponenten sind für das Lenken eines Glycosylierungs-Enzyms zu einem geeigneten Ort verantwortlich.
- Lokalisierungs-Sequenzen von einem Organismus können auch in anderen Organismen funktionieren. Von der Membran-durchdringenden Region einer α-2,6-Sialyltransferase (α-2,6-ST) aus Ratten zum Beispiel, ein Enzym, von dem man weiß, dass es in dem Trans-Golgi der Ratte lokalisiert ist, wurde gezeigt, dass es auch für die Lokalisierung eines Reportergens (Invertase) in dem Hefe-Golgi verantwortlich ist (Schwientek, et al., 1995). Genau die gleiche Membran-durchdringende Region wurde jedoch als Teil einer α-2,6-Sialyltransferase von vollständiger Länge in dem ER zurückgehalten und nicht weiter in den Golgi von Hefe transportiert (Krezdorn et al., 1994). Ein menschliches GalT von vollständiger Länge wurde trotz nachgewiesener hoher Transkriptionsspiegel in Hefe nicht einmal synthetisiert. Andererseits war die Transmembran-Region des gleichen menschlichen GalT, nachdem sie an einen Invertase-Reporter fusioniert worden war, in der Lage die Lokalisation hin zum Hefe-Golgi auszurichten, wenn auch in niedrigen Produktionsspiegeln. Schwientek und Mitarbeiter zeigten, dass die Fusion von 28 Aminosäuren einer Hefe-Mannosyltransferase (Mnt1), ein Bereich, umfassend einen N-terminalen cytoplasmatischen Schwanz, eine Transmembran-Region und acht Aminosäuren der Stamm-Region, mit der katalytischen Domäne von menschlichem GalT für die Lokalisation eines aktiven GalT im Golgi ausreichend ist (Schwientek et al. 1995 J. Biol. Chem. 270 (10): 5483–5489). Andere Galactosyltransferasen scheinen sich auf Wechselwirkungen mit Enzymen zu stützen, die in bestimmten Organellen ansässig sind, da sie nach Entfernen ihrer Transmembran-Region noch zur richtigen Lokalisation in der Lage sind.
- Die falsche Lokalisation eines Glycosylierungs-Enzyms kann das richtige Funktionieren des Enzyms im Stoffwechselweg verhindern. Aspergillus nidulans, zum Beispiel, der zahlreiche α-1,2-Mannosidasen besitzt (Eades and Hintz, 2000 Gene 255 (1): 25–34), fügt kein GlcNAc an Man5GlcNAc2 an, wenn er mit dem Kaninchen GnTI-Gen transformiert wurde, trotz eines hohen allgemeinen Spiegels an GnTI-Aktivität (Kalsner et al., 1995). GnTI, obwohl in aktiver Form exprimiert, kann unkorrekt lokalisiert sein, derart, dass das Enzym mit beiden seiner Substrate keinen Kontakt bekommt: das im Entstehen befindliche N-Glycan des Glycoproteins und UDP-GlcNAc. Alternativ kann es möglich sein, dass der Wirtsorganismus die angemessenen Spiegel an UDP-GlcNAc in dem Golgi nicht bereitstellt.
- Glycoproteine, die therapeutisch verwendet werden
- Ein wesentlicher Anteil der Proteine, die aus Menschen oder anderen Tieren isoliert werden, sind glycosyliert. Unter den Proteinen, die therapeutisch verwendet werden, sind etwa 70% glycosyliert. Wenn jedoch ein therapeutisches Protein in einem Mikroorganismus wie Hefe hergestellt wird und unter Verwendung des endogenen Stoffwechselwegs glycosyliert wird, ist seine therapeutische Wirksamkeit im Allgemeinen stark reduziert. Solche Glycoproteine sind im Allgemeinen in Menschen immunogen und zeigen nach der Verabreichung eine verkürzte in vivo-Halbwertszeit (Takeuchi, 1997).
- Spezifische Rezeptoren in Menschen und Tieren können terminale Mannose-Reste erkennen und die rasche Beseitigung des Proteins aus dem Blutstrom verstärken. Zusätzliche nachteilige Wirkungen können Veränderungen in der Faltung, der Löslichkeit, der Zugänglichkeit für Proteasen, dem Verkehr, dem Transport, der Kompartimentierung, der Sekretion, der Erkennung durch andere Proteine oder Faktoren, der Antigenität oder der Allergenität des Proteins beinhalten. Entsprechend war es notwendig therapeutische Glycoproteine in tierischen Wirtsystemen herzustellen, derart, dass das Muster an Glycosylierung identisch oder zumindest ähnlich dem in Menschen oder zu den beabsichtigten Empfänger-Arten ist. In den meisten Fällen wird ein Säuger-Wirtsystem wie die Säuger-Zellkultur verwendet.
- Systeme für die Herstellung therapeutischer Glycoproteine
- Um therapeutische Proteine herzustellen, die geeignete Glycoformen haben und zufriedenstellende therapeutische Wirkungen besitzen, sind Expressionssysteme verwendet worden, die auf Tieren oder Pflanzen beruhen. Die erhältlichen Systeme beinhalten:
- 1. Ovar-Zellen des chinesischen Hamsters (CHO), Maus-Fibroblasten-Zellen und Maus-Myolom-Zellen (Arzneimittelforschung. 1998 Aug; 48 (8): 870–880);
- 2. transgene Tiere wie Ziegen, Schafe, Mäuse und andere (Dente Prog. Clin. Biol. 1989 Res. 300: 85–98, Ruther et al., 1988 Cell 53 (6): 847–856; Ware, J., et al. 1993 Thrombosis and Haemostasis 69 (6): 1194–1194; Cole, E. S., et al. 1994 J. Cell Biochem. 265–265);
- 3. Pflanzen (Arabidopsis thaliana, Tabak etc.) (Staub, et al. 2000 Nature Biotechnology 18 (3): 333–338) (McGarvey, P. B., et al. 1995 Bio-Technology 13 (13): 1484–1487; Bardor, M., et al. 1999 Trends in Plant Science 4 (9): 376–380);
- 4. Insekten-Zellen (Spodoptera frugiperda Sf9, Sf21, Trichoplusia ni, etc. in Kombination mit rekombinanten Baculoviren, wie multiples nukleäres Polyeder-Virus von Autographa californica (das Lepidoptera-Zellen infiziert) (Altmans et al., 1999 Glycoconj. J. 16 (2): 109–123).
- Rekombinante menschliche Proteine, die in den vorstehend erwähnten Wirtsystemen exprimiert werden, können noch nicht-menschliche Glycoformen beinhalten (Raju et al., 2000 Annals Biochem. 283 (2): 123–132). Im Besonderen kann dem Anteil der N-Glycane die terminale Sialinsäure fehlen, die im Allgemeinen in menschlichen Glycoproteinen gefunden wird. Wesentliche Anstrengungen waren auf die Entwicklung von Verfahren zum Erhalt von Glycoproteinen ausgerichtet, die den menschlichen Formen hinsichtlich der Struktur so nahe wie möglich sind oder die andere therapeutische Vorteile besitzen. Glycoproteine, die spezifische Glycoformen besitzen, können besonders zweckdienlich sein, zum Beispiel für die zielgerichtete Lenkung therapeutischer Proteine. So kann zum Beispiel das Anfügen von einem oder mehreren Sialinsäure-Resten an eine Glycan-Seitenkette die Lebensdauer eines therapeutischen Glycoproteins nach Verabreichung in vivo erhöhen. Demgemäß können Säuger-Wirtzellen gentechnisch verändert sein, um den Umfang an terminalen Sialinsäuren in Glycoproteinen zu erhöhen, die in den Zellen exprimiert sind. Alternativ dazu kann Sialinsäure vor der Verabreichung in vitro an das Protein von Interesse konjugiert werden, wobei eine Sialinsäuretransferase und ein geeignetes Substrat verwendet werden. Zusätzlich wurden Veränderungen in der Zusammensetzung des Wachstums-Mediums oder der Expression von Enzymen, die an der menschlichen Glycosylierung beteiligt sind, verwendet, um Glycoproteine herzustellen, die den menschlichen Formen ähnlicher sind (S. Weikert, et al., Nature Biotechnology, 1999, 17, 1116–1121; Werner, Noe, et al. 1998 Arzneimittelforschung 48 (8): 870–880; Weikert, Papac et al., 1999; Andersen and Goochee 1994 Cur. Opin. Biotechnol. 5: 546–549; Yang and Butler 2000 Biotechnol. Bioengin. 68 (4): 370–380). Alternativ dazu können gezüchtete menschliche Zellen verwendet werden.
- Alle die bestehenden Systeme haben jedoch signifikante Nachteile. Nur bestimmte therapeutische Proteine sind für die Expression in Säuger- oder Pflanzen-Systemen geeignet (z.B. solche, denen jegliche cytotoxische Wirkung fehlt oder eine andere Wirkung, die für Wachstum nachteilig ist). Tierische und Pflanzen-Zellkultur-Systeme sind gewöhnlich sehr langsam, benötigen häufig mehr als eine Woche Wachstum unter sorgfältig kontrollierten Bedingungen, um jede beliebige zweckmäßige Menge des Proteins von Interesse herzustellen. Im Vergleich mit denjenigen aus mikrobiellen Fermentations-Verfahren ist der Ertrag an Proteinen trotzdem ungünstig. Zusätzlich benötigen Zellkultur-Systeme im Allgemeinen komplexe und aufwendige Nährstoffe und Co-Faktoren wie fötales Kälberserum. Darüber hinaus kann das Wachstum durch programmierten Zelltod (Apoptose) eingeschränkt sein.
- Darüber hinaus sind tierische Zellen (insbesondere Säuger-Zellen) in hohem Maße empfänglich für virale Infektion oder Kontamination. In einigen Fällen kann das Virus oder andere infektiöse Mittel das Wachstum der Kultur beeinträchtigen, während in anderen Fällen das Mittel ein menschliches Pathogen sein kann, das das therapeutische Protein-Produkt hinsichtlich seiner beabsichtigten Verwendung unbrauchbar macht. Darüber hinaus benötigen zahlreiche Zellkultur-Verfahren die Verwendung von komplexen, temperatursensitiven, aus Tieren stammenden Wachstumsmedium-Bestandteile, die Pathogene tragen können, wie bovine spongiforme Enzephalopathie (BSE)-Prionen. Solche Pathogene sind schwierig nachzuweisen und/oder schwierig zu entfernen oder zu sterilisieren, ohne das Wachstumsmedium zu beeinträchtigen. In jedem Fall macht die Verwendung von tierischen Zellen zur Herstellung therapeutischer Proteine kostspielige Qualitätskontrollen erforderlich, um die Produkt-Sicherheit zu gewährleisten.
- Zur Herstellung großer Volumina therapeutischer Proteine wie menschliches Serum-Albumin, Gewebe-Plasminogen-Aktivator, monoclonale Antikörper, Hämoglobin, Kollagen, Fibrinogen und andere können auch transgene Tiere verwendet werden. Während transgene Ziegen und andere transgene Tiere (Mäuse, Schafe, Kühe, etc.) gentechnisch verändert werden können, sodass sie therapeutische Proteine in hohen Konzentrationen in der Milch produzieren, ist das Verfahren kostspielig, da jede Charge sich einer drastischen Qualitätskontrolle unterziehen muss. Tiere können eine Vielzahl- tierischer oder menschlicher Pathogene tragen, einschließlich Bakterien, Viren, Pilze und Prionen. Im Falle der Traberkrankheit und der bovinen spongiformen Enzephalopathie kann das Testen, um eine Infektion auszuschließen, etwa ein Jahr dauern. Die Herstellung von therapeutischen Verbindungen wird somit vorzugsweise in einer gut kontrollierten sterilen Umgebung durchgeführt, z.B. unter Bedingungen Guter Herstellerischer Praxis (GMP). Es ist jedoch im Allgemeinen nicht durchführbar, Tiere in solchen Umgebungen zu halten. Darüber hinaus beruht transgene Tier-Technologie auf verschiedenen Tieren und ist somit von Natur aus nicht einheitlich, wohingegen Zellen, die in einem Fermenter wachsen, von einer gut charakterisierten Original-Zellbank (MCB) abstammen. Darüber hinaus können externe Faktoren wie unterschiedliche Nahrungsaufnahme, Krankheit und mangelnde Homogenität innerhalb einer Herde das Glycosylierungs-Muster des Endproduktes beeinflussen. Es ist zum Beispiel in Menschen bekannt, dass verschiedene Ernährungs-Angewohnheiten in verschiedenen Glycosylierungs-Mustern resultieren.
- Transgene Pflanzen sind als eine potenzielle Quelle entwickelt worden, um Proteine von therapeutischem Wert zu erhalten. Die Expression von Proteinen in hohen Mengen in Pflanzen leidet jedoch unter dem Gene Silencing, einem Mechanismus, durch den die Gene für stark exprimierte Proteine in der nachfolgenden Pflanzen-Generation herunterreguliert sind. Zusätzlich fügen Pflanzen Xylose und/oder α-1,3-gekoppelte Fucose an N-Glycane von Proteinen an, was in Glycoproteinen resultiert, die sich in der Struktur von tierischen unterscheiden und in Säugern immunogen sind (Atlmann, Marz et al., 1995 Glycoconj. J. 12 (2); 150–155). Darüber hinaus ist es im Allgemeinen nicht durchführbar Pflanzen in einer sterilen oder GMP-Umgebung zu züchten und die Gewinnung von Proteinen aus Pflanzen-Geweben ist kostspieliger als die Gewinnung aus fermentierten Mikroorganismen. Glycoprotein-Herstellung, wobei eukaryontische Mikroorganismen verwendet werden Der Mangel an einem geeigneten Expressionssystem ist somit ein signifikantes Hindernis für die kostengünstige und sichere Herstellung von rekombinanten menschlichen Glycoproteinen. Die Herstellung von Glycoproteinen durch die Fermentation von Mikroorganismen würde zahlreiche Vorteile gegenüber den bestehenden Systemen bieten. Auf Fermentation basierende Verfahren können zum Beispiel Folgendes bieten; (a) die schnelle Herstellung von hohen Konzentrationen an Protein; (b) die Möglichkeit sterile, gut kontrollierte Herstellungsbedingungen zu verwenden (z.B. GMP-Bedingungen); (c) die Möglichkeit einfache, chemisch definierte Wachstumsmedien zu verwenden; (d) die Leichtigkeit genetischer Manipulation; (e) das Fehlen von kontaminierenden menschlichen oder tierischen Pathogenen; (f) die Möglichkeit eine große Bandbreite an Proteinen zu exprimieren, einschließlich diejenigen, die in Folge von Toxizität etc. in der Zellkultur schlecht exprimiert werden; (g) die Leichtigkeit der Protein-Gewinnung (z.B. über Sekretion in das Medium). Zusätzlich sind Fermentations- Anlagen im Allgemeinen wesentlich weniger kostspielig zu konstruieren als Zellkultur-Anlagen.
- Wie vorstehend bemerkt, glycosylieren Bakterien, einschließlich Arten wie Escherichia coli, die im Allgemeinen verwendet werden, um rekombinante Proteine herzustellen, Proteine jedoch nicht in einer spezifischen Art und Weise wie Eukaryonten. Verschiedene methylotrophe Hefen wie Pichia pastoris, Pichia methanolica und Hansenula polymorpha sind besonders zweckmäßig als eukaryontische Expressionssysteme, da sie in der Lage sind zu hohen Zelldichten zu wachsen und/oder große Mengen an rekombinantem Protein zu sekretieren. Wie vorstehend bemerkt, unterscheiden sich jedoch Glycoproteine, die in diesen eukaryontischen Mikroorganismen exprimiert werden, wesentlich in der N-Glycan-Struktur von denjenigen in Tieren. Dies hat die Verwendung von Hefe oder filamentösen Pilzen als Wirte für die Herstellung von zahlreichen zweckmäßigen Glycoproteinen verhindert.
- Einige Anstrengungen sind unternommen worden, um die Glycosylierungswege von eukaryontischen Mikroorganismen zu modifizieren, um Glycoproteine bereitzustellen, die für die Verwendung als therapeutische Mittel in Säugern besser geeignet sind. Es sind zum Beispiel einige Glycosyltransferasen vereinzelt cloniert und in S. cerevisiae (GalT, GnT I), Aspergillus nidulans (GnT I) und anderen Pilzen (Yoshida et al., 1999, Kalsner et al., 1995 Glycoconj. J. 12(3): 360–370, Schwientek et al., 1995) cloniert worden. Es wurden jedoch keine N-Glycane mit menschlichen Eigenschaften erhalten.
- Hefen produzieren eine Vielzahl von Mannosyltransferasen, z.B. 1,3-Mannosyltransferasen (z.B. MNN1 in S. cerevisiae) (Graham and Emr, 1991 J. Cell Biol. 114 (2): 207–218), 1,2-Mannosyltransferasen (z.B. die KTR/KRE-Familie von S. cerevisiae), 1,6-Mannosyltransferasen (OCH1 von S. cerevisiae), Mannosylphosphat-Transferasen (MNN4 und MNN6 von S. cerevisiae) und zusätzliche Enzyme, die an endogenen Glycosylierungs-Reaktionen beteiligt sind. Viele dieser Gene sind vereinzelt deletiert worden, wobei lebensfähige Organismen entstanden sind, die veränderte Glycosylierungs-Profile besitzen. Beispiele sind in Tabelle 1 gezeigt.
- Tabelle 1. Beispiele von Hefe-Stämmen, die eine veränderte Mannosylierung besitzen

- Zusätzlich offenbart die Japanische Patent-Anmeldung mit der Veröffentlichungs-Nr. 8-336387 einen mutierten OCH1-Stamm von Pichia pastoris. Das OCH1-Gen codiert 1,6-Mannosyltransferase, die eine Mannose an die Glycan-Struktur Man8GlcNAc2 anfügt, wobei Man9GlcNAc2 erhalten wird. Die Man9GlcNAc2-Struktur ist dann ein Substrat für weitere Mannosylierung in vivo, was dann zu den hypermannosylierten Glycoproteinen führt, die für Hefen charakteristisch sind und im Allgemeinen mindestens 30 bis 40 Mannose-Reste pro N-Glycan haben können. In dem mutierten OCH1-Stamm sind Proteine angereichert, die mit Man8GlcNAc2 glycosyliert sind, und Hypermannosylierung tritt nicht auf. Die Struktur Man8GlcNAc2 ist jedoch kein Substrat für tierische Glycosylierungs-Enzyme wie menschliche UDP-GlcNAc-Transferase I und entsprechend ist das Verfahren für die Herstellung von Proteinen mit menschlichen Glycosylierungs-Mustern nicht zweckmäßig.
- Martinet und Mitarbeiter (Biotechnol. Lett. 1998, 20 (12), 1171–1177) berichteten über die Expression einer α-1,2-Mannosidase von Trichoderma reesei in P. pastoris. Es wurde einiges Mannosetrimmen der N-Glycane eines Modell-Proteins beobachtet. Das Modell-Protein hatte jedoch keine N-Glycane mit der Struktur Man5GlcNAc2, was als ein Zwischenprodukt für die Erzeugung von komplexen N-Glycanen notwendig wäre. Demgemäß ist das Verfahren nicht zweckmäßig für die Herstellung von Proteinen mit menschlichen oder tierischen Glycosylierungs-Mustern.
- In ähnlicher Weise exprimierten Chiba und Mitarbeiter 1998 α-1,2-Mannosidase von Aspergillus saitoi in der Hefe Saccharomyces cerevisiae. Eine Signalpeptid-Sequenz (His-Asp-Glu-Leu) wurde gentechnisch in die exogene Mannosidase rekombiniert, um das Zurückhalten in dem endoplasmatischen Retikulum zu verstärken. Zusätzlich handelte es sich bei dem Hefe-Wirt um eine Mutante, der drei Enzym-Aktivitäten fehlten, die mit Hypermannosylierung von Proteinen assoziiert sind: 1,6-Mannosyltransferase (OCH1); 1,3-Mannosyltransferase (MNN1); und Mannosylphosphat-Transferase (MNN4). Die N-Glycane des dreifach mutierten Wirtes bestanden somit aus der Struktur Man8GlcNAc2, die gegenüber den High-Mannose-Formen bevorzugt wird, die in Wildtyp-S. cerevisiae gefunden werden. In Anwesenheit der stark exprimierten gentechnisch veränderten Mannosidase wurden die N-Glycane eines Modellproteins (Carboxypeptidase Y) ausgetrimmt, wobei ein Gemisch, bestehend aus 27 Mol% Man5GlcNAc2, 22 Mol% Man6GlcNAc2, 22 Mol% Man7GlcNAc2 und 29 Mol% Man8GlcNAc2 erhalten wurde. Das Trimmen der endogenen Zellwand-Glycoproteine war weniger effizient, nur 10 Mol% der N-Glycane hatten die gewünschte Man5GlcNAc2-Struktur. Über Chibas Arbeit ist auch in EP-A1 1211311 berichtet worden. Obwohl Chiba eine mehr als ausreichende Menge seiner Mannosidase in hoher Kopienzahl exprimierte und er relativ geringfügige Erträge erhielt, schlägt er vor, dass sogar noch mehr Enzym exprimiert werden sollte, um das Problem der niedrigen Erträge auszuräumen. Er schlug niemals vor, dass das gewünschte Enzym so ausgewählt werden sollte, dass es in der ausgewählten Wirtszelle an dem subzellulären Ort, an dem es sich befindet, optimal arbeitet, um sein Substrat umzuwandeln.
- Da ausschließlich die Man5GlcNAc2-Glycane für eine weitere enzymatische Umwandlung zu menschlichen Glycoformen zugänglich sind, ist das Verfahren nicht wirksam für die Herstellung von Proteinen, die menschliche Glycosylierungs-Muster besitzen. In Proteinen, die eine einzige N-Glycosylierungsstelle besitzen, hätten mindestens 73 Mol% eine unkorrekte Struktur. In Proteinen, die zwei bzw. drei N-Glycosylierungsstellen besitzen, hätten mindestens 93 bzw. 98 Mol% eine unkorrekte Struktur. Solche niedrigen Wirkungsgrade bezüglich der Umwandlung sind nicht befriedigend für die Herstellung von therapeutischen Mitteln, insbesondere da die Abtrennung von Proteinen, die verschiedene Glycoformen besitzen, im Allgemeinen kostspielig und schwierig ist.
- Mit dem Ziel, ein mehr menschenähnliches Glycoprotein bereitzustellen, das von einem Pilz als Wirt abstammt, offenbart das US-Patent Nr. 5,834,251 von Maras und Contreras ein Verfahren zur Herstellung eines Hybrid-Glycoproteins, das von Trichoderma reesei abstammt. Ein Hybrid-N-Glycan hat an dem Man-α1-6-Arm des Kerns ausschließlich Mannose-Reste und eine oder zwei komplexe Antennen an dem Man-α1-3-Arm. Während diese Struktur Nutzen hat, hat das Verfahren den Nachteil, dass zahlreiche enzymatische Schritte in vitro durchgeführt werden müssen, was kostspielig und zeitraubend ist. Isolierte Enzyme sind teuer herzustellen und zu erhalten, können ungewöhnliche und kostspielige Substrate benötigen (z.B. UDP-GlcNAc) und neigen unter den verwendeten Bedingungen zu Verlust an Aktivität und/oder zu Proteolyse.
- Deshalb ist es ein Ziel der vorliegenden Erfindung ein System und Verfahren für humanisierende Glycosylierung von rekombinanten Glycoproteinen bereitzustellen, die in Pichia pastoris und anderen niederen Eukaryonten wie Hansenula polymorpha, Pichia stiptis, Pichia methanolica, Pichia sp, Kluyveromyces sp, Candida albicans, Aspergillus nidulans und Trichoderma reesei exprimiert werden.
- ZUSAMMENFASSUNG DER ERFINDUNG
- Es wurden Zelllinien entwickelt, die gentechnisch modifizierte Glycosylierungswege besitzen, die es ihnen erlauben eine Folge von enzymatischen Reaktionen durchzuführen, die die Prozessierung von Glycoproteinen in Menschen nachahmen. Rekombinante Proteine, die in diesen gentechnisch veränderten Wirten exprimiert werden, stellen Glycoproteine bereit, die ihren menschlichen Gegenstücken ähnlicher, wenn nicht im Wesentlichen mit diesen identisch sind. Die niederen Eukaryonten, die im Allgemeinen High-Mannose-enthaltende N-Glycane produzieren, einschließlich unizelluläre und multizelluläre Pilze wie Pichia pastoris, Hansenula polymorpha, Pichia stiptis, Pichia methanolica, Pichia sp., Klyveromyces sp., Candida albicans, Aspergillus nidulans und Trichoderma reesei werden modifiziert, sodass sie N-Glycane wie Man5GlcNAc2 oder andere Strukturen entlang des menschlichen Glycosylierungswegs produzieren. Dies wird erreicht, indem eine Kombination von gentechnischen Verfahren und/oder einer Auswahl von Stämmen verwendet wird, die: keine bestimmten Enzyme exprimieren, die die unerwünschten komplexen Strukturen erzeugen, die für die Glycoproteine von Pilzen charakteristisch sind, die exogene Enzyme exprimieren, die so ausgewählt wurden, dass sie entweder unter den Bedingungen, die in dem Pilz vorliegen, wo Aktivität erwünscht ist, optimale Aktivität besitzen oder die auf ein Organell gerichtet sind, wo optimale Aktivität erreicht wird, und Kombinationen daraus, in denen der genetisch veränderte Eukaryont multiple exogene Enzyme exprimiert, die erforderlich sind, um "Menschen-ähnliche"-Glycoproteine zu produzieren.
- In einer ersten Ausführungsform betrifft die Erfindung eine Wirtszelle, die ein unizellulärer oder filamentöser Pilz ist, der keine α-1,6-Mannosyltransferase-Aktivität, bezogen auf das N-Glycan auf einem Glycoprotein, aufweist, und die in ihrem endoplasmatischen Retikulum (ER) oder Golgi-Apparat ein Hybridenzym hat, das ausgewählt ist, um optimale Aktivität im ER oder Golgi der Wirtszelle zu haben, sodass die Wirtszelle 50 bis 100 Mol% Man5GlcNAc2 auf einem Substrat-Glycoprotein bilden kann, wobei das Hybridenzym umfasst:
- (a) eine exogene Mannosidase-katalytische Domäne mit optimaler Aktivität im ER oder Golgi bei einem pH-Wert zwischen 5,1 oder 8,0; verbunden mit
- (b) einem zellulären Zielsignal-Peptid, das normalerweise nicht mit der katalytischen Domäne von (a) verbunden ist, wobei das zelluläre Zielsignal-Peptid die exogene Mannosidase-katalytische Domäne in das ER oder den Golgi-Apparat bringt.
- Somit betrifft die Erfindung einen Mikroorganismus, der gentechnisch verändert ist, um ein exogenes α-1,2-Mannosidase-Enzym zu exprimieren, das einen optimalen pH-Wert zwischen 5,1 und 8,0 besitzt, vorzugsweise zwischen 5,9 und 7,5. Somit ist das exogene Enzym auf das endoplasmatische Retikulum oder den Golgi-Apparat des Wirtsorganismus gerichtet, wo es N-Glycane wie Man8GlcNAc2 derart trimmt, dass Man5GlcNAc2 erhalten wird. Die letztere Struktur ist zweckmäßig, weil sie mit einer Struktur übereinstimmt, die in Säugern, insbesondere Menschen, gebildet wird; sie ist ein Substrat für weitere Glycosylierungs-Reaktionen in vivo und/oder in vitro, die ein endgültiges N-Glycan produzieren, das dem ähnlich oder zu dem identisch ist, das in Säugern, insbesondere in Menschen gebildet wird; und sie ist kein Substrat für Hypermannosylierungs-Reaktionen, die in vivo in Hefe und anderen Mikroorganismen auftreten und die ein Glycoprotein hochgradig immunogen in Säugern machen.
- In einer zweiten Ausführungsform betrifft die Erfindung ein Verfahren zur Herstellung der Wirtszelle, die hierin offenbart ist, wobei das Verfahren den Schritt umfasst, diesen unizellulären oder filamentösen Pilz mit einem Nucleinsäure-Molekül zu transformieren, das ein Hybridenzym codiert, das ausgewählt ist, um optimale Aktivität im ER oder Golgi der Wirtszelle zu haben, sodass die Wirtszelle 50 bis 100 Mol% Man5GlcNAc2 auf einem Substrat-Glycoprotein bilden kann, wobei das Hybridenzym umfasst:
- (a) eine exogene Mannosidase-katalytische Domäne mit optimaler Aktivität im ER oder Golgi bei einem pH-Wert zwischen 5,1 und 8,0; verbunden mit
- (b) einem zellulären Zielsignal-Peptid, das normalerweise nicht mit der katalytischen Domäne von (a) verbunden ist, wobei das zelluläre Ziesignal-Peptid die exogene katalytische Domäne in das ER oder den Golgi-Apparat bringt.
- In einer dritten Ausführungsform betrifft die Erfindung ein Verfahren zur Herstellung eines Glycoproteins in einer Wirtszelle, die ein unizellulärer oder filamentöser Pilz ist, der keine α-1,6-Mannosyltransferase-Aktivität in Bezug auf das N-Glycan auf einem Glycoprotein aufweist, wobei das Verfahren die Schritte umfasst:
- (a) Exprimieren des Glycoproteins in einer Population von Wirtszellen, die mit einer DNA-Bibliothek transformiert sind, die mindestens zwei verschiedene genetische Konstrukte umfasst, wobei mindestens eines davon ein DNA-Fragment umfasst, das ein Hybridenzym codiert, umfassend:
- (i) eine Glycosylierungsenzym-katalytische Domäne mit optimaler Aktivität im ER oder Golgi bei einem pH-Wert zwischen 5,1 und 8,0; verbunden mit
- (ii) einem zellulären Zielsignal-Peptid, das normalerweise nicht mit der katalytischen Domäne von (i) verbunden ist, wobei das zelluläre Zielsignal-Peptid auf das ER oder den Golgi-Apparat der Wirtszelle abzielt; und
- (b) Auswählen der Wirtszelle, die ein Glycoprotein mit einem gewünschten Glycosylierungs-Muster auf dem Glycoprotein zeigt.
- Somit ist der Glycosylierungs-Signalweg eines eukaryontischen Mikroorganismus in dieser Ausführungsform modifziert, indem (a) eine DNA-Bibliothek erzeugt wird, die mindestens zwei Gene beinhaltet, die exogene Glycosylierungs-Enzyme codieren; (b) indem der Mikroorganismus mit der Bibliothek transformiert wird, um eine genetisch gemischte Population zu erzeugen, die mindestens zwei voneinander verschiedene exogene Glycosylierungs-Enzyme exprimiert; (c) indem aus der Population ein Mikroorganismus ausgewählt wird, der den gewünschten Glycosylierungs-Phänotyp besitzt. In einer anderen Ausführungsform beinhaltet eine DNA-Bibliothek chimäre Gene, wobei jedes eine Protein-Lokalisierungs-Sequenz und eine katalytische Aktivität codiert, die mit Glycosylierung in Beziehung steht. Entsprechend stellt die Erfindung in einer vierten Ausführungsform eine DNA-Bibliothek bereit, die genetische Konstrukte umfasst, wobei die Konstrukte ein DNA-Fragment umfassen, das ein zelluläres Zielsignal-Peptid codiert, das auf das endoplasmatische Retikulum oder den Golgi-Apparat abzielt, das im Leserahmen mit einem DNA-Fragment ligiert ist, das ein Glycosylierungs-Enzym oder ein katalytisch aktives Fragment davon codiert, das in dem ER oder Golgi bei einem pH-Wert zwischen 5,1 und 8,0 optimale Aktivität besitzt, wobei die DNA-Bibliothek mindestens zwei DNA-Fragmente umfasst, die ein zelluläres Zielsignal-Peptid codieren, und mindestens zwei DNA-Fragmente umfasst, die exogene Glycosylierungs-Enzyme codieren. Organismen, die unter Verwendung dieses Verfahrens modifiziert wurden, sind zweckmäßig zur Herstellung von Glycoproteinen, die ein Glycosylierungs-Muster aufweisen, das dem von Säugern, insbesondere Menschen, ähnlich oder dazu identisch ist.
- In einer fünften Ausführungsform ist der Glycosylierungs-Signalweg modifiziert, sodass er ein Zucker-Nucleotid-Transport-Enzym exprimiert. In einer bevorzugten Ausführungsform wird auch ein Nucleotid-Diphosphatase-Enzym exprimiert. Der Transporter und die Diphosphatase verbessern die Wirksamkeit der gentechnisch veränderten Glycosylierungsschritte, indem die geeigneten Substrate für die Glycosylierungs-Enzyme in den geeigneten Kompartimenten bereitgestellt werden, die kompetitive Produkthemmung reduziert wird und das Entfernen von Nucleosid-Diphosphaten verstärkt wird.
- Schließlich betrifft die Erfindung eine Glycoprotein-Zusammensetzung, die durch die hierin offenbarten Verfahren erhältlich ist und die 50 bis 100 Mol% von Man5GlcNAc2 umfasst, das in vivo durch GnTI zu GlcNAcMan5GlcNAc2 umgewandelt wird, wobei diesem Glycoprotein Galactose, Fucose und terminale Sialinsäure fehlen.
- BESCHREIBUNG DER FIGUREN
-
1A ist ein schematisches Diagramm des typischen Pilz-N-Glycosylierungs-Signalwegs. -
1B ist ein schematisches Diagramm eines typischen menschlichen N-Glycosylierungs-Signalwegs. - AUSFÜHRLICHE BESCHREIBUNG DER ERFINDUNG
- Die Verfahren und rekombinanten niederen Eukaryonten-Stämme, die hierin beschrieben sind, werden verwendet, um "humanisierte Glycoproteine" herzustellen. Die rekombinanten niederen Eukaryonten werden hergestellt, indem niedere Eukaryonten, die eines oder mehrere Enzyme, die bei der Herstellung von High-Mannose-Strukturen beteiligt sind, nicht exprimieren, derart gentechnisch verändert werden, dass sie die Enzyme exprimieren, die erforderlich sind, um Menschenähnliche Zucker zu produzieren. Wie hierin verwendet, ist ein niederer Eukaryont ein einzelliger oder filamentöser Pilz. Wie hierin verwendet, bezieht sich ein "humanisiertes Glycoprotein" auf ein Protein, an das N-Glycane angeheftet sind, die weniger als vier Mannose-Reste beinhalten, und die synthetischen Zwischenprodukte (die ebenfalls zweckmäßig sind und in vitro weiter manipuliert werden können), die mindestens fünf Mannose-Reste besitzen. In einer bevorzugten Ausführungsform enthalten die Glycoprotein, die in dem rekombinanten niederen eukaryontischen Stamm produziert werden, mindestens 27 Mol% des Man5-Zwischenprodukts. Dies wird erreicht, indem eine bessere Mannosidase cloniert wird, d.h. ein Enzym, das so ausgewählt wird, dass es unter den Bedingungen, die in dem Organismus vorliegen, an der Stelle, an der die Proteine glycosyliert werden, optimale Aktivität besitzt oder durch Lenken des Enzyms zu dem Organell, wo die Aktivität gewünscht ist.
- In einer bevorzugten Ausführungsform werden eukaryontische Stämme verwendet, die ein oder mehrere Enzyme nicht exprimieren, die an der Produktion von High-Mannose-Strukturen beteiligt sind. Diese Stämme können durch gentechnische Manipulation erzeugt werden oder einer von diesen zahlreichen Mutanten sein, die bereits in Hefen beschrieben sind, einschließlich einer Hypermannosylierungs-minus(OCH1)-Mutante in Pichia pastoris.
- Die Stämme können für jeweils ein einzelnes Enzym gentechnisch verändert werden oder es kann eine Bibliothek von Genen erzeugt werden, die potenziell zweckmäßige Enzyme codiert, und diejenigen Stämme können ausgewählt werden, die Enzyme mit optimalen Aktivitäten besitzen oder die die am meisten "Menschenähnlichen" Glycoproteine produzieren.
- Niedere Eukaryonten, die in der Lage sind Glycoproteine herzustellen, die das angeheftete N-Glycan Man5GlcNAc2 besitzen, sind besonders zweckmäßig, denn, da (a) ihnen eine hochgradige Mannosylierung fehlt (z.B. mehr als acht Mannosen pro N-Glycan oder im Besonderen 30–40 Mannosen), zeigen sie reduzierte Immunogenität in Menschen; und (b) das N-Glycan ist ein Substrat für weitere Glycosylierungs-Reaktionen, wobei eine sogar noch mehr Menschen-ähnliche Glycoform gebildet wird, z.B. durch die Wirkung von GlcNAc-Transferase I, wobei GlcNAcMan5GlcNAc2 gebildet wird. Man5GlcNAc2 muss in vivo in einem hohen Ertrag gebildet werden, zumindest vorübergehend, da alle nachfolgenden Glycosylierungs-Reaktionen Man5GlcNAc2 oder ein Derivat davon benötigen. Entsprechend wird ein Ertrag von mehr als 27 Mol%, mehr bevorzugt ein Ertrag von 50–100 Mol% Glycoproteine erhalten, in denen ein hoher Anteil von N-Glycanen Man5GlcNAc2 besitzen. Bevorzugte erfindungsgemäße Wirtszellen erbringen 50–100 Mol% Glycoproteine, in denen N-Glycane Man5GlcNAc2 besitzen, das in vivo durch GlcNAc-Transferase I umgewandelt wurde. Es ist dann möglich in vitro weitere Glycosylierungs-Reaktionen durchzuführen, wobei zum Beispiel das Verfahren des US-Patents Nr. 5,834,251 von Maras und Contreras verwendet wird. In einer bevorzugten Ausführungsform wird mindestens eine weitere Glycosylierungs-Reaktion in vivo durchgeführt. In einer hochgradig bevorzugten Ausführungsform davon werden aktive Formen weiterer Glycosylierungs-Enzyme in dem endoplasmatischen Retikulum und/oder dem Golgi-Apparat exprimiert.
- Wirts-Mikroorganismen
- Hefe und filamentöse Pilze sind beide für die Herstellung von rekombinanten Proteinen, sowohl intrazelluläre als auch sekretierte, erfolgreich verwendet worden (Cereghino, J. L. and J. M. Cregg 2000 FEMS Microbiology Reviews 24 (1): 45–66; Harkki, A., et al. 1989 Bio-Technology 7 (6): 596; Berka, R. M., et al. 1992 Abstr. Papers Amer. Chem. Soc. 203: 121-BIOT; Svetina, M., et al. 2000 J. Biotechnol. 76 (2–3): 245–251.
- Obwohl die Glycosylierung in Hefe und Pilzen sich von der in Menschen stark unterscheidet, werden einige Elemente gemeinsam benutzt. Der erste Schritt, der Transfer der Kern-Oligosaccharid-Struktur an das nascierende Protein ist in allen Eukaryonten, einschließlich Hefen, Pilze, Pflanzen und Menschen, hoch konserviert (vergleiche
1A und1B ). Die nachfolgende Prozessierung des Kern-Oligosaccharids unterscheidet sich jedoch signifikant in Hefe und beinhaltet das Anhängen von einigen Mannose-Zuckern. Dieser Schritt wird durch Mannosyltransferasen katalysiert, die sich in dem Golgi befinden (z.B. OCH1, MNT1, MNN1, etc.), die sequenziell Mannose-Zucker an das Kern-Oligosaccharid anheften. Die resultierende Struktur ist für die Herstellung Menschen-ähnlicher Proteine unerwünscht und somit ist es wünschenswert, die Mannosyltransferase-Aktivität zu reduzieren oder auszuschalten. Für Mutanten von S. cerevisiae, die für Mannosyltransferase-Aktivität defizient sind (z.B. och1- oder mnn9-Mutanten), konnte gezeigt werden, dass sie nicht letal sind und in dem Oligosaccharid von Hefe-Glycoproteinen einen reduzierten Mannose-Gehalt zeigen. In Abhängigkeit von dem jeweiligen endogenen Glycosylierungs-Muster des Wirts kann es auch erforderlich sein, dass andere Oligosaccharid-prozessierende Enzyme wie Mannosylphosphat-Transferase ebenfalls ausgeschaltet werden müssen. Nach der Reduktion unerwünschter endogener Glycosylierungs-Reaktionen muss die Bildung von komplexen N-Glycanen das Wirtssystem eingebracht werden. Dies erfordert die stabile Expression von einigen Enzymen und Zucker-Nucleotid-Transportern. Darüber hinaus müssen diese Enzyme auf eine Art und Weise lokalisiert werden, dass eine sequenzielle Prozessierung der reifenden Glycosylierungs-Struktur gewährleistet ist. - Ziel-Glycoproteine
- Die hierin beschriebenen Verfahren sind für die Herstellung von Glycoproteinen zweckmäßig, insbesondere Glycoproteine, die therapeutisch in Menschen verwendet werden. Solche therapeutischen Proteine werden typischerweise durch Injektion, oral, pulmonal oder auf andere Art und Weise verabreicht.
- Beispiele für geeingete Ziel-Glycoproteine beinhalten, ohne Einschränkung: Erythropoietin, Cytokine wie Interferon-α, Interferon-β, Interferon-γ, Interferon-ω und Granulocyten-CSF, Koagulationsfaktoren wie Faktor VIII, Faktor IX und menschliches Protein C, lösliche IgE-Rezeptor-α-Kette, IgG, IgM, Urokinase, Chymase und Harnstoff-Trypsin-Inhibitor, IGF-bindendes Protein, epidermaler Wachstumsfaktor, Wachstumshormon-feisetzender Faktor, Annexin V-Fusionsprotein, Angiostatin, vaskulärer endothelialer Wachstumsfaktor-2, myeloider Vorläufer-Inhibitor-Faktor-1 und Osteoprotegerin.
- Verfahren zur Herstellung von Glycoproteinen, die das N-Glycan Man5GlcNAc2 umfassen
- Der erste Schritt beinhaltet die Auswahl oder die Erzeugung eines niederen Eukaryonten, der in der Lage ist eine spezifische Vorläufer-Struktur von Man5GlcNAc2 zu produzieren, die in der Lage ist in vivo durch die Wirkung einer GlcNAc-Transferase I GlcNAc aufzunehmen. Dieser Schritt erfordert die Bildung einer bestimmten isomeren Struktur von Man5GlcNAc2. Diese Struktur muss innerhalb der Zelle in einem hohen Ertrag gebildet werden (mehr als 30%), da alle nachfolgenden Manipulationen von der Anwesenheit dieser Vorstufe abhängig sind. Man5GlcNAc2-Strukturen sind für die Bildung von komplexem N-Glycan erforderlich, ihr Vorliegen ist jedoch in keinster Weise ausreichend, da Man5GlcNAc2 in verschiedenen isomeren Formen auftreten kann, die als ein Substrat für GlcNAc-Transferase I dienen können oder nicht. Die meisten Glycosylierungs-Reaktionen sind nicht vollständig und somit enthält ein bestimmtes Protein im Allgemeinen eine Auswahl unterschiedlicher Kohlenhydrat-Strukturen (d.h. Glycoformen) auf seiner Oberfläche. Die bloße Anwesenheit von Spuren-Mengen (weniger als 5%) einer bestimmten Struktur wie Man5GlcNAc2, ist von geringer praktischer Relevanz. Es ist die Bildung eines bestimmten GLcNAc-Transferase I akzeptierenden Zwischenprodukts (Struktur I) in hohem Ertrag (oberhalb von 30%), die erforderlich ist. Die Bildung dieses Zwischenprodukts ist notwendig und erlaubt im Nachfolgenden die in vivo-Synthese von komplexen N-Glycanen.
- Um die Struktur in vivo bereitzustellen, kann man solche niederen Eukaryonten aus der Natur auswählen oder alternativ dazu vorliegende Pilze oder andere niedere Eukaryonten gentechnisch manipulieren. Es wurde bisher kein niederer Eukaryont gezeigt, der solche Strukturen in vivo in höherem Maße als 1,8% in Bezug auf die gesamten N-Glycane bereitstellt (Mayas et al., 1997), sodass ein gentechnisch manipulierter Organismus bevorzugt wird. Verfahren wie diejenigen, die im US-Patent Nr. 5,595,900 beschrieben sind, können verwendet werden, um die Abwesenheit oder Anwesenheit von bestimmten Glycosyltransferasen, Mannosidasen und Zucker-Nucleotid-Transportern in einem Zielorganismus von Interesse zu identifizieren.
- Inaktivierung von Pilz-Glycosylierungs-Enzymen wie 1,6-Mannosyltransferase
- Das hierin beschriebene Verfahren kann verwendet werden, um das Glycosylierungs-Muster einer großen Bandbreite niederer Eukaryonten gentechnisch zu manipulieren (z.B. Hansenula polymorpha, Pichia stiptis, Pichia methanolica, Pichia sp, Kluyveromyces sp, Candida albicans, Aspergillus nidulans, Trichoderma reesei etc.). Pichia pastoris wird verwendet, um die erforderlichen Manipulationsschritte beispielhaft darzustellen. Ähnlich wie andere niedere Eukaryonten prozessiert P. pastoris Man>9GlcNAc2-Strukturen in dem ER mit einer 1,2-α-Mannosidase, wobei Man8GlcNAc2 erhalten wird. Durch die Wirkung einiger Mannosyltransferasen wird diese Struktur dann in hypermannosylierte Strukturen umgewandelt (Man9GlcNAc2), die auch als Mannane bekannt sind. Zusätzlich wurde gefunden, dass P. pastoris in der Lage ist nicht-terminale Phophatgruppen über die Wirkung von Mannosylphosphat-Transferasen an die Kohlenhydrat-Struktur anzuhängen. Das ist im Gegensatz zu den Umsetzungen, die in Säugerzellen gefunden werden, die das Entfernen von Mannose-Zucker beinhalten, im Gegensatz zu deren Hinzufügung. Es ist von besonderer Wichtigkeit die Fähigkeit des Pilzes auszuschalten, die vorliegende Man8GlcNAc2-Struktur zu hypermannolysieren. Das kann erreicht werden, indem entweder ein Pilz ausgewählt wird, der nicht hypermannosyliert ist oder indem ein solcher Pilz gentechnisch manipuliert wird.
- Gene, die an diesem Prozess beteiligt sind, wurden in Pichia pastoris identifiziert und durch Erzeugen von Mutationen in diesen Genen ist man in der Lage die Herstellung von "unerwünschten" Glycoformen zu reduzieren. Solche Gene können durch Homologie zu bestehenden Mannosyltransferasen (z.B. OCH1, MNN4, MNN6, MNN1) identifiziert werden, die in anderen niederen Eukaryonten wie C. albicans, Pichia angusta oder S. cerevisiae gefunden werden, oder indem der Wirtsstamm mutiert wird und auf einen Phänotyp mit reduzierter Mannosylierung selektiert wird. Basierend auf Homologien zwischen bekannten Mannosyltransferasen und Mannosylphosphat-Transferasen kann man entweder PCR-Primer entwerfen, wofür Beispiele in Tabelle 2 gezeigt sind, oder Gene oder Genfragmente, die solche Enzyme codieren, als Sonden verwenden, um Homologe in den DNA-Bibliotheken der Ziel-Organismen zu identifizieren. Alternativ dazu kann es möglich sein bestimmte Phänotypen in verwandten Organismen zu ergänzen. Um zum Beispiel das Gen oder die Gene zu erhalten, die 1,6-Mannosyltransferase-Aktivität in P. pastoris codieren, würde man die nachfolgenden Schritte durchführen. OCH1-Mutanten von S. cerevisiae sind temperatursensitiv und wachsen langsam bei erhöhten Temperaturen. Somit kann man funktionelle Homologe von OCH1 in P. pastoris identifizieren, indem man eine OCH1-Mutante von S. cerevisiae mit einer DNA- oder cDNA-Bibliothek aus P. pastoris ergänzt. Solche Mutanten von S. cerevisiea können auf der Seite http://genome-www.stanford.edu/Saccharomyces/ gefunden werden und sind auf der Seite http://www.resgen.com/products/YEASTD.php3 kommerziell erhältlich. Mutanten, die bei erhöhter Temperatur einen Phänotyp mit normalem Wachstum zeigen, nachdem sie mit einer P. pastoris-DNA-Bibliothek transformiert worden waren, tragen wahrscheinlich ein OCH1-Homologes von P. pastoris. Eine solche Bibliothek kann erzeugt werden, indem chromosomale DNA von P. pastoris mit einem geeigneten Restriktionsenzym partiell gespalten wird und die gespaltene DNA nach Inaktivierung des Restriktionsenzyms in einen geeigneten Vektor ligiert wird, der mit einem passenden Restriktionsenzym gespalten worden war. Geeignete Vektoren sind pRS314, ein Plasmid mit niedriger Kopienzahl (CEN6/ARS4), das auf pBluescript basiert, das den Trp1-Marker enthält (Sikorski, R. S., and Hieter, P., 1989, Genetics 122, S. 19–27), oder pFL44S, ein Plasmid von hoher Kopienzahl (2μ), das auf einem modifizierten pUC19 basiert, das den URA3-Marker enthält (Bonneaud, N., et al., 1991, Yeast 7, S. 609–615). Solche Vektoren werden gewöhnlich von akademischen Wissenschaftlern verwendet oder ähnliche Vektoren sind von einer Vielzahl unterschiedlicher Anbieter erhältlich, wie Invitrogen (Carlsbad, CA), Pharmacia (Piscataway, NJ), New England Biolabs (Beverly, MA). Beispiele sind pYES/GS, 2μ-Replikationsursprung, basierend auf dem Hefe-Expressionsplasmid von Invitrogen, oder Yep24, ein Clonierungs-Vehikel von New England Biolabs. Nach Ligierung der chromosomalen DNA und des Vektors kann man die DNA-Bibliothek in einen Stamm von S. cerevisiae mit einer spezifischen Mutation transformieren und hinsichtlich der Korrektur des entsprechenden Phänotyps selektionieren. Nach Subclonierung und Sequenzierung des DNA-Fragments, das in der Lage ist den Wildtyp-Phänotyp wieder herzustellen, kann man dieses Fragment verwenden, um die Aktivität des Genprodukts auszuschalten, das in P. pastoris von OCH1 codiert wird.
- Falls die gesamte genomische Sequenz eines bestimmten Pilzes von Interesse bekannt ist, kann man alternativ solche Gene identifizieren, indem man einfach öffentlich erhältliche DNA-Datenbanken durchsucht, die von einigen Quellen erhältlich sind, wie NCBI, Swissprot etc. Indem man zum Beispiel eine vorgegebene genomische Sequenz oder eine Datenbank mit einem bekannten 1,6-Mannosyltransferase-Gen (OCH1) aus S. cerevisiae durchsucht, kann man in der Lage sein, in einem solchen Genom Gene mit hoher Homologie zu identifizieren, die mit einem hohen Grad an Sicherheit ein Gen codieren, das 1,6-Mannosyltransferase-Aktivität besitzt. Unter Verwendung jedes dieser beiden Ansätze wurden Homologe zu einigen bekannten Mannosyltransferasen von S. cerevisiae in P. pastoris identifiziert. Diese Gene besitzen ähnliche Funktionen wie diejenigen Gene, die bei der Mannosylierung von Proteinen in S. cerevisiae beteiligt sind, und somit kann ihre Deletion dazu verwendet werden das Glycosylierungs-Muster in P. pastoris oder einem beliebigen anderen Pilz mit ähnlichen Glycosylierungs-Signalwegen zu manipulieren.
- Die Erzeugung von Gen-knock outs, nachdem eine vorgegebene Ziel-Gensequenz bestimmt worden ist, ist im Bereich der Hefe- und Pilzmolekularbiologie ein gut etabliertes Verfahren und kann von jedem beliebigen durchschnittlichen Fachmann durchgeführt werden (R. Rothsteins, (1991) Methods in Enzymology, Bd, 194, S. 281). In der Tat kann die Wahl eines Wirts-Organismus durch die Verfügbarkeit von guten Transformations- und Gen-Zerstörungs-Verfahren für einen solchen Wirt beeinflusst werden. Falls einige Mannosyltransferasen einem Knock-out ausgesetzt werden müssen, erlaubt das Verfahren, das von Alani und Kleckner entwickelt wurde, die wiederholte Verwendung der URA3-Marker, um nacheinander sämtliche unerwünschte endogene Mannosyltransferase-Aktivität auszuschalten. Dieses Verfahren wurde durch andere verfeinert, beinhaltet aber im Grundsatz die Verwendung von zwei sich wiederholenden DNA-Sequenzen, die einen gegen selektierbaren Marker flankieren. Zum Beispiel: URA3 kann als ein Marker verwendet werden, um die Selektion einer Transformante zu gewährleisten, die ein Konstrukt integriert hat. Durch Flankierung des URA3-Markers mit direkten sich wiederholenden Sequenzen, kann man als erstes nach Transformanten selektionieren, die das Konstrukt integriert haben und somit das Zielgen zerstört haben. Nach Isolierung der Transformanten und deren Charakterisierung kann man in einer zweiten Runde nach denjenigen gegenselektionieren, die gegen 5' FOA resistent sind. Kolonien, die in der Lage sind, auf Platten zu überleben, die 5' FOA enthalten, haben den URA3-Marker durch ein Crossing over-Ereignis, an dem die vorstehend erwähnten sich wiederholenden Sequenzen beteiligt sind, wieder verloren. Dieser Ansatz erlaubt somit die wiederholte Verwendung des gleichen Markers und erleichtert die Zerstörung von multiplen Genen, ohne zusätzliche Marker zu benötigen.
- Das Ausschalten spezifischer Mannosyltransferasen wie 1,6-Mannosyltransferase (OCH1), Mannosylphosphat-Transferasen (MNN4, MNN6 oder Gene, die Ibd-Mutanten komplementieren) in P. pastoris erlaubt die Erzeugung von gentechnisch veränderten Stämmen dieses Organismus, die vorrangig Man8GlcNAc2 synthetisieren und somit verwendet werden können, um das Glycosylierungs-Muster weiter zu modifizieren, um komplexeren menschlichen Glycoform-Strukturen in stärkerem Maße ähnlich zu sein. Eine bevorzugte Ausführungsform dieses Verfahrens verwendet bekannte DNA-Sequenzen, die bekannte biochemische Glycosylierungs-Aktivitäten codieren, um ähnliche oder identische biochemische Funktionen in P. pastoris auszuschalten, sodass die Glycosylierungs-Struktur des resultierenden gentechnisch veränderten P. pastoris-Stammes modifiziert ist. Tabelle 2.Legende: M = A oder C, R = A oder G, W = A oder T, S = C oder G, Y = C oder T, K = G oder T, V = A oder C oder G, H = A oder C oder T, D = A oder G oder T, B = C oder G oder T, N = G oder A oder T oder C.

- Einfügung einer Mannosidase in den gentechnisch veränderten Wirt
- Das hierin beschriebene Verfahren ermöglicht es eine solche Struktur mit hohem Ertrag zu erhalten, für den Zweck, diese zu modifizieren, um komplexe N-Glycane zu erhalten. Ein erfolgreiches Schema, um geeignete Man5GlcNAc2-Strukturen zu erhalten, muss zwei parallel laufende Ansätze beinhalten: (1) Das Verringern der endogenen Mannosyltransferase-Aktivität und (2) das Entfernen von 1,2-α-Mannose durch Mannosidasen, um hohe Spiegel an geeigneten Man5GlcNAc2-Strukturen zu erhalten. Was dieses Verfahren von dem Stand der Technik unterscheidet ist, dass es diese beiden Probleme direkt bewältigt. Wie die Arbeit von Chiba und Mitarbeitern zeigt, kann man Man8GlcNAc2-Strukturen in S. cerevisiae in ein Man5GlcNAc2-Isomer reduzieren, indem man das Vorliegen einer Pilz-Mannosidase von A. Saitoi durch gentechnische Verfahren in das große ER einbringt. Die Mängel von deren Verfahren sind zweifach: (1) In der extrazellulären Glycoprotein-Fraktion werden keine ausreichenden Mengen von Man5GlcNAc2 gebildet (10%), und (2) es ist nicht klar, dass die in vivo gebildete Man5GlcNAc2-Struktur in der Tat in der Lage ist, GlcNAc durch die Wirkung von GlcNAc-Transferase zu akzeptieren. Falls einige Glycosylierungsstellen in einem gewünschten Protein vorliegen, ergibt sich die Wahrscheinlichkeit (P), ein solches Protein in einer korrekten Form zu erhalten, aus der Beziehung P = (F)n, wobei n der Anzahl an Glycosylierungsstellen entspricht und F der Fraktion an gewünschten Glycoformen entspricht. Ein Glycoprotein mit drei Glycosylierungsstellen würde eine Wahrscheinlichkeit von 0,1% besitzen, die geeigneten Vorstufen für das Prozenieren von komplexen und Hybrid-N-Glycanen an allen Glycosylierungsstellen bereitzustellen, was den wirtschaftlichen Wert eines solchen Ansatzes beschränkt.
- Die meisten Enzyme, die in dem ER und dem Golgi-Apparat von S. cerevisiae aktiv sind, haben ein pH-Optimum, das zwischen 6,5 und 7,5 liegt (siehe Tabelle 3). Alle vorherigen Ansätze, die Mannosylierung durch die Wirkung von rekombinanten Mannosidasen zu reduzieren, haben sich auf Enzyme konzentriert, die ein pH-Optimum um etwa pH 5,0 besitzen (Martinet et al., 1998, und Chiba et al., 1998), obwohl die Aktivität dieser Enzyme bei pH 7,0 sogar auf weniger als 10% reduziert ist und diese somit höchstwahrscheinlich an dem Ort ihrer Verwendung, dem ER und dem frühen Golgi von P. pastoris und S. cerevisiae, nicht ausreichende Aktivität bereitstellen. Ein bevorzugtes Verfahren verwendet eine α-Mannosidase in vivo, wobei das pH-Optimum der Mannosidase nicht mehr als 1,4 pH-Einheiten vom durchschnittlichen pH-Optimum anderer repräsentativer Marker-Enzyme abweicht, die in dem/den gleichen Organellen) lokalisiert sind. Das pH-Optimum des Enzyms, das auf ein spezifischen Organell abzielt, sollte mit dem pH-Optimum von anderen Enzymen übereinstimmen, die in dem gleichen Organell gefunden werden, sodass die maximale Aktivität pro Einheit Enzym erhalten wird. Tabelle 3 fasst die Aktivität von Mannosidasen aus verschiedenen Quellen und deren jeweilige pH-Optima zusammen. Tabelle 4 fasst deren Lokalisation zusammen.
- Tabelle 3. Mannosidasen und ihr pH-Optimum

- Wenn man versucht High-Mannose-Strukturen zu trimmen, um Man5GlcNAc2 in dem ER oder dem Golgi-Apparat von S. cerevisiae zu erhalten, kann man ein beliebiges Enzym oder eine Kombination von Enzymen auswählen, das/die (1) ein ausreichend enges pH-Optimum hat haben (d.h. zwischen pH 5,2 und pH 7,8), und (2) von dem/denen man weiß, dass es/sie alleine oder gemeinsam die spezifische isomere Man5GlcNAc2-Struktur erzeugt/erzeugen, die erforderlich ist, um das nachfolgende Hinzufügen von GlcNAc durch GnTI zu gewährleisten. Jedes beliebige Enzym oder jede beliebige Kombination von Enzymen, das/die gezeigt hat/haben, dass es/sie eine Struktur erzeugt/erzeugen, die in vitro durch GnTI in GlcNAcMan5GlcNAc2 umgewandelt werden kann, würde eine geeignete Wahl darstellen. Diese Kenntnis kann aus der wissenschaftlichen Literatur erhalten werden oder experimentell durch Bestimmen, dass eine potenzielle Mannosidase Man8GlcNAc2-PA in Man5GlcNAc2-PA umwandeln kann, und dann Testen, ob die erhaltene Man5GlcNAc2-PA-Struktur als ein Substrat für GnTI und UDP-GlcNAc dienen kann, um GlcNAcMan5GlcNAc2 in vitro zu erhalten. Mannosidase IA von einer menschlichen oder murinen Quelle würde zum Beispiel eine geeignete Wahl sein.
- 1,2-Mannosidase-Aktivität in dem ER und dem Golgi
- Vorherige Versuche Mannosylierung durch die Wirkung von clonierten exogenen Mannosidasen zu reduzieren sind daran gescheitert Glycoproteine bereitzustellen, die eine ausreichende Fraktion (z.B. > 27 Mol%) an N-Glycanen enthalten, die die Struktur Man5GlcNAc2 besitzen (Martinet et al., 1998, und Chiba et al., 1998). Diese Enzyme sollten effizient in dem ER oder dem Golgi-Apparat funktionieren, um effizient nascierende Glycoproteine umzuwandeln. Während die beiden Mannosidasen, die im Stand der Technik verwendet werden (aus A. saitoi und T. reesei), ein pH-Optimum von 5,0 besitzen, haben die meisten Enzyme, die in dem ER und dem Golgi-Apparat von Hefe (z.B. S. cerevisiae) aktiv sind, pH-Optima, die zwischen 6,5 und 7,5 liegen (siehe Tabelle 3). Da die Glycosylierung von Proteinen ein hoch entwickeltes und wirkungsvolles Verfahren darstellt, kann geschlossen werden, dass der interne pH-Wert des ER und des Golgi ebenfalls in dem Bereich von etwa 6–8 liegt. Bei pH 7,0 ist die Aktivität der Mannosidasen, die im Stand der Technik verwendet werden, auf weniger als 10% reduziert, was für die wirkungsvolle Man5GlcNAc2-Produktion in vivo nicht ausreichend ist. Chibas Arbeit wurde auch in EP-A1 1 211 310 berichtet. Weitere Anmerkungen sind hierin vorstehend bereitgestellt.
- Tabelle 4. Zelluläre Lokalisation und pH-Optima von verschiedenen mit Glycosylierung in Zusammenhang stehenden Enzymen von S. cerevisiae.

- Das α-1,2-Mannosidase-Enzym sollte bei einem pH-Wert zwischen 5,1 und 8,0 optimale Aktivität besitzen. In einer bevorzugten Ausführungsform hat das Enzym eine optimale Aktivität bei einem pH-Wert zwischen 5,9 und 7,5. Der optimale pH-Wert kann unter in vitro-Test-Bedingungen bestimmt werden. Bevorzugte Mannosidasen beinhalten diejenigen, die in Tabelle 3 aufgelistet sind und die geeignete pH-Optima besitzen, z.B. Aspergillus nidulans, Homo sapiens IA (Golgi), Homo sapiens IB (Golgi), Lepidoptera-Insekten-Zellen (IPLB-SF21AE), Homo sapiens, Maus-IB (Golgi) und Xanthomonas manihotis. In einer bevorzugten Ausführungsform wird ein einzelnes cloniertes Mannosidase-Gen in dem Wirtsorganismus exprimiert. Es kann jedoch in einigen Fällen wünschenswert sein einige verschiedene Mannosidase-Gene zu exprimieren oder einige Kopien eines bestimmten Gens, um die angemessene Herstellung von Man5GlcNAc2 zu erreichen. In den Fällen, in denen multiple Gene verwendet werden, sollten die codierten Mannosidasen alle pH-Optima innerhalb des bevorzugten Bereiches von 5,1 bis 8,0 besitzen oder insbesondere zwischen 5,9 und 7,5. In einer speziell bevorzugten Ausführungsform wird die Mannosidase-Aktivität zu dem ER oder den cis-Golgi geleitet, wo die frühen Reaktionen der Glycosylierung auftreten.
- Bildung von komplexen N-Glycanen
- Ein zweiter Schritt des Verfahrens beinhaltet das aufeinanderfolgende Anhängen von Zuckern an die nascierende Kohlenhydrat-Struktur durch Einbringen der Expression von Glucosyltransferasen in den Golgi-Apparat. Dieser Prozess benötigt als erstes die funktionelle Expression von GnTI in dem frühen oder medialen Golgi-Apparat sowie die Gewährleistung der Bereitstellung von UDP-GlcNAc in ausreichenden Mengen.
- Integrations-Stellen
- Da das ultimative Ziel dieses gentechnischen Manipulationsversuchs ein robuster Stamm für die Herstellung von Protein ist, der in der Lage ist dies in einem industriellen Fermentations-Verfahren gut durchzuführen, beinhaltet die Integration multipler Gene in das Pilz-Chromosom eine sorgfältige Planung. Der veränderte Stamm wird sehr wahrscheinlich mit einer Auswahl unterschiedlicher Gene transformiert werden müssen und diese Gene werden in einer stabilen Art und Weise transformiert werden müssen, um zu gewährleisten, dass die gewünschte Aktivität während des Fermentations-Prozesses erhalten bleibt. Jede beliebige Kombination der nachfolgenden Enzym-Aktivitäten wird in den Proteinexpressions-Pilzwirt eingebracht werden müssen: Sialyltransferasen, Mannosidasen, Fucosyltransferasen, Galactosyltransferasen, Glucosyltransferasen, GlcNAc-Transferasen, ER- und Golgi-spezifische Transporter (z.B. Syn- und Antiport-Transporter für UDP-Galactose und andere Vorstufen), andere Enzyme, die an der Prozessierung von Oligosacchariden beteiligt sind, und Enzyme, die bei der Synthese von aktivierten Oligosaccharid-Vorstufen beteiligt sind, wie UDP-Galactose, CMP-N-Acetyl-Neuraminsäure. Gleichzeitig wird eine Auswahl von Genen ausgeschaltet werden müssen, die Enzyme codieren, von denen bekannt ist, dass sie für nicht-menschliche-Glycosylierungs-Reaktionen charakteristisch sind.
- Leiten von Glycosyltransferasen zu spezifischen Organellen:
- Glycosyltransferasen und Mannosidasen kleiden die innere (luminale) Oberfläche des ER und des Golgi-Apparates aus und stellen dabei eine "katalytische" Oberfläche bereit, die die aufeinanderfolgende Prozessierung von Glycoproteinen erlaubt, während diese durch das ER und das Golgi-Netzwerk voranschreiten. In der Tat stellen die multiplen Kompartimente des Cis-, des medialen und des Trans-Golgi und das Trans-Golgi-Netzwerk (TGN) die veschiedenen Orte bereit, in denen die geordnete Aufeinanderfolge von Glycosylierungs-Reaktionen stattfinden kann. Während ein Glycoprotein von der Synthese in dem ER zu seiner vollen Reife in dem späten Golgi oder dem TGN voranschreitet, ist es in aufeinanderfolgenden Schritten veschiedenen Glycosidasen, Mannosidasen und Glycosyl-Transferasen ausgesetzt, sodass eine spezifische Kohlenhydrat-Struktur synthetisiert werden kann. Viel Arbeit wurde aufgewendet, um den genauen Mechanismus zu offenbaren, durch den diese Enzyme in ihren jeweiligen Organellen festgehalten und verankert werden. Das sich entwickelnde Bild ist komplex, aber Hinweise legen nahe, dass die Stamm-Region, die Membrandurchdringende Region und der cytoplasmatische Schwanz einzeln oder gemeinsam die Enzyme an die Membran von einzelnen Organellen dirigieren und dabei die assoziierte katalytische Domäne an diesen Ort lokalisieren.
- Zielsequenzen sind gut bekannt und in der wissenschaftlichen Literatur und in öffentlichen Datenbanken beschrieben, wie nachfolgend im Hinblick auf Bibliotheken zur Auswahl von Zielsequenzen und zielgerichteten Enzymen ausführlicher diskutiert.
- Verfahren zur Herstellung einer Bibliothek, um modifizierte Glycosylierungswege zu produzieren.
- Eine Bibliothek, die mindestens zwei Gene beinhaltet, die exogene Glycosylierungs-Enzyme codieren, wird in den Wirtsorganismus transformiert, wobei eine genetisch gemischte Population hergestellt wird. Transformanten, die den gewünschten Glycosylierungs-Phänotyp besitzen, werden dann aus der gemischten Population selektioniert. In einer bevorzugten Ausführungsform ist der Wirtsorganismus eine Hefe, insbesondere P. pastoris, und der Wirts-Glycosylierungsweg wird durch die funktionelle Expression von einem oder mehreren menschlichen oder tierischen Glycosylierungs-Enzymen modifiziert, wobei Protein-N-Glycane erhalten werden, die menschlichen Glycoformen ähnlich oder mit diesen identisch sind. In einer speziell bevorzugten Ausführungsform beinhaltet die DNA-Bibliothek genetische Konstrukte, die Fusionen von Glycosylierungs-Enzymen mit Zielsequenzen für verschiedene zelluläre Loci codieren, die an der Glycosylierung beteiligt sind, insbesondere das ER, den Cis-Golgi, den medialen Golgi oder den Trans-Golgi.
- Beispiele von Modifikationen für Glycosylierung, die unter Verwendung des Verfahrens durchgeführt werden können, sind: (1) die Veränderung eines eukaryontischen Mikroorganismus derart, dass er Mannose-Reste von Man8GlcNAc2 trimmt, sodass Man5GlcNAc2 als ein Protein-N-Glycan entsteht; (2) Veränderung eines eukaryontischen Mikroorganismus derart, dass durch die Wirkung von GlcNAc- Transferase I ein N-Acetyl-Glucosamin (GlcNAc)-Rest an Man5GlcNAc2 angehängt wird; (3) Veränderung eines eukaryontischen Mikroorganismus derart, dass er funktionell ein Enzym wie eine N-Acetyl-Glucosamin-Transferase (GnTI, GnTII, GnTIII, GnTIV, GnTV, GnTVI), eine Mannosidase II, eine Fucosyltransferase, eine Galactosyltransferase (GalT) oder Sialyltransferasen (ST) exprimiert.
- Durch Wiederholen des Verfahrens können zunehmend komplexe Glycosylierungswege in den Ziel-Mikroorganismus eingebracht werden. In einer bevorzugten Ausführungsform wird der Wirtsorganismus zwei oder mehrere Male mit DNA-Bibliotheken transformiert, die Sequenzen enthalten, die Glycosylierungs-Aktivitäten codieren. Nach jeder Runde der Transformation oder in einer alternativen Ausführungsform nachdem einige Transformationen aufgetreten sind, kann die Selektionierung der gewünschten Phänotypen durchgeführt werden. Auf diese Art und Weise können komplexe Glycosylierungswege schnell eingefügt werden.
- DNA-Bibliotheken
- Es ist erforderlich eine DNA-Bibliothek zusammenzustellen, die mindestens zwei exogene Gene beinhaltet, die Glycosylierungs-Enzyme codieren. Zusätzlich zu den Sequenzen im offenen Leserahmen wird es im Allgemeinen bevorzugt jedes Konstrukt der Bibliothek mit solchen Promotoren, Transkriptions-Terminatoren, Enhancern, Ribosomen-Bindungsstellen und anderen funktionellen Sequenzen bereitzustellen, wie es notwendig ist, um nach Transformation in den Wirtsorganismus eine effektive Transkription und Translation der Gene zu gewährleisten. Dort, wo der Wirt Pichia pastoris ist, beinhalten geeignete Promotoren zum Beispiel die AOX1-, AOX2-, DAS- und P40-Promotoren. Es wird auch bevorzugt, jedes Konstrukt mit mindestens einem selektionierbaren Marker bereitzustellen, wie ein Gen, das eine Arzneistoff-Resistenz verleiht oder das einen Defekt im Wirts-Metabolismus komplementiert. Das Vorliegen des Markers ist für die nachfolgende Selektion von Transformanten zweckmäßig; in Hefe können zum Beispiel die URA3-, HIS4-, SUC2-, G418-, BLA- oder SH BLE-Gene verwendet werden.
- In einigen Fällen kann die Bibliothek direkt aus existierenden oder Wildtyp-Genen zusammengesetzt werden. In einer bevorzugten Ausführungsform wird die DNA-Bibliothek jedoch aus der Fusion von zwei oder mehreren Unter-Bibliotheken zusammengesetzt. Durch die Ligierung der Unter-Bibliotheken im Leserahmen ist es möglich eine große Anzahl von neuen genetischen Konstrukten zu erzeugen, die zweckmäßige zielgerichtete Glycosylierungs-Aktivitäten codieren. Eine zweckmäßige Unter-Bibliothek beinhaltet zum Beispiel DNA-Sequenzen, die irgendeine beliebige Kombination von Enzymen wie Sialyltransferasen, Mannosidasen, Fucosyltransferasen, Galactosyltransferasen, Glucosyltransferasen und GlcNAc-Transferasen codieren. Vorzugsweise sind die Enzyme von menschlichem Ursprung, obwohl andere Säuger-, tierische oder Pilz-Enzyme ebenfalls zweckmäßig sind. In einer bevorzugten Ausführungsform sind die Gene verkürzt, wobei Fragmente erhalten werden, die die katalytischen Domänen der Enzyme codieren. Durch das Entfernen der endogenen Ziel-Sequenzen können die Enzyme dann von neuem ausgerichtet und an anderen zellulären Orten exprimiert werden. Die Auswahl solcher katalytischer Domänen kann durch die Kenntnis der jeweiligen Umgebung geleitet werden, in der die katalytische Domäne im Nachfolgenden aktiv sein soll. Wenn ein bestimmtes Glycosylierungs-Enzym zum Beispiel in dem späten Golgi aktiv sein soll und alle bekannten Enzyme des Wirtsorganismus in dem späten Golgi ein bestimmtes pH-Optimum besitzen, dann wird eine katalytische Domäne ausgewählt, die bei diesem pH-Wert eine angemessene Aktivität besitzt.
- Eine andere zweckmäßige Unter-Bibliothek beinhaltet DNA-Sequenzen, die Signalpeptide codieren, die in der Lokalisation eines Proteins an einen bestimmten Ort innerhalb des ER, des Golgi oder des Trans-Golgi-Netzwerks resultieren. Diese Signalsequenzen können aus dem Wirtsorganismus sowie von anderen verwandten oder nicht verwandten Organismen ausgewählt werden. Membran-gebundene Proteine des ER oder des Golgi können typischerweise zum Beispiel N-terminale Sequenzen beinhalten, die einen cytosolischen Schwanz (ct), eine Transmembran-Domäne (tmd) und eine Stamm-Region (sr) codieren. Die ct-, tmd- und sr-Sequenzen sind ausreichend, einzeln oder in Kombination, um Proteine an die innere (luminale) Membran des Organells zu verankern. Entsprechend beinhaltet eine bevorzugte Ausführungsform der Unter-Bibliothek von Signalsequenzen ct-, tmd- und/oder sr-Sequenzen dieser Proteine. In einigen Fällen ist es wünschenswert die Unter-Bibliothek mit unterschiedlichen Längen der sr-Sequenz bereitzustellen. Dies kann durch PCR erreicht werden, wobei Primer verwendet werden, die an das 5'-Ende der DNA binden, die die cytosolische Region codiert, und wobei eine Serie von gegenläufigen Primern verwendet wird, die an verschiedene Stellen der Stammregion binden. Noch andere zweckmäßige Quellen von Signalsequenzen beinhalten Wiederfindungs-Signalpeptide, z.B. die Tetrapeptide HDEL oder KDEL, die im Allgemeinen an dem C-terminalen Ende von Proteinen gefunden werden, die retrograd in das ER oder den Golgi transportiert werden. Noch andere Quellen von Signalsequenzen beinhalten (a) Typ II-Membranproteine, (b) die Enzyme, die in Tabelle 3 aufgelistet sind, (c) Membran-durchdringende Nucleotid-Zucker-Transporter, die in dem Golgi lokalisiert sind, und (d) Sequenzen, auf die sich in Tabelle 5 bezogen wird.
- Tabelle 5. Quellen zweckmäßiger Zielsequenzen für Kompartimente
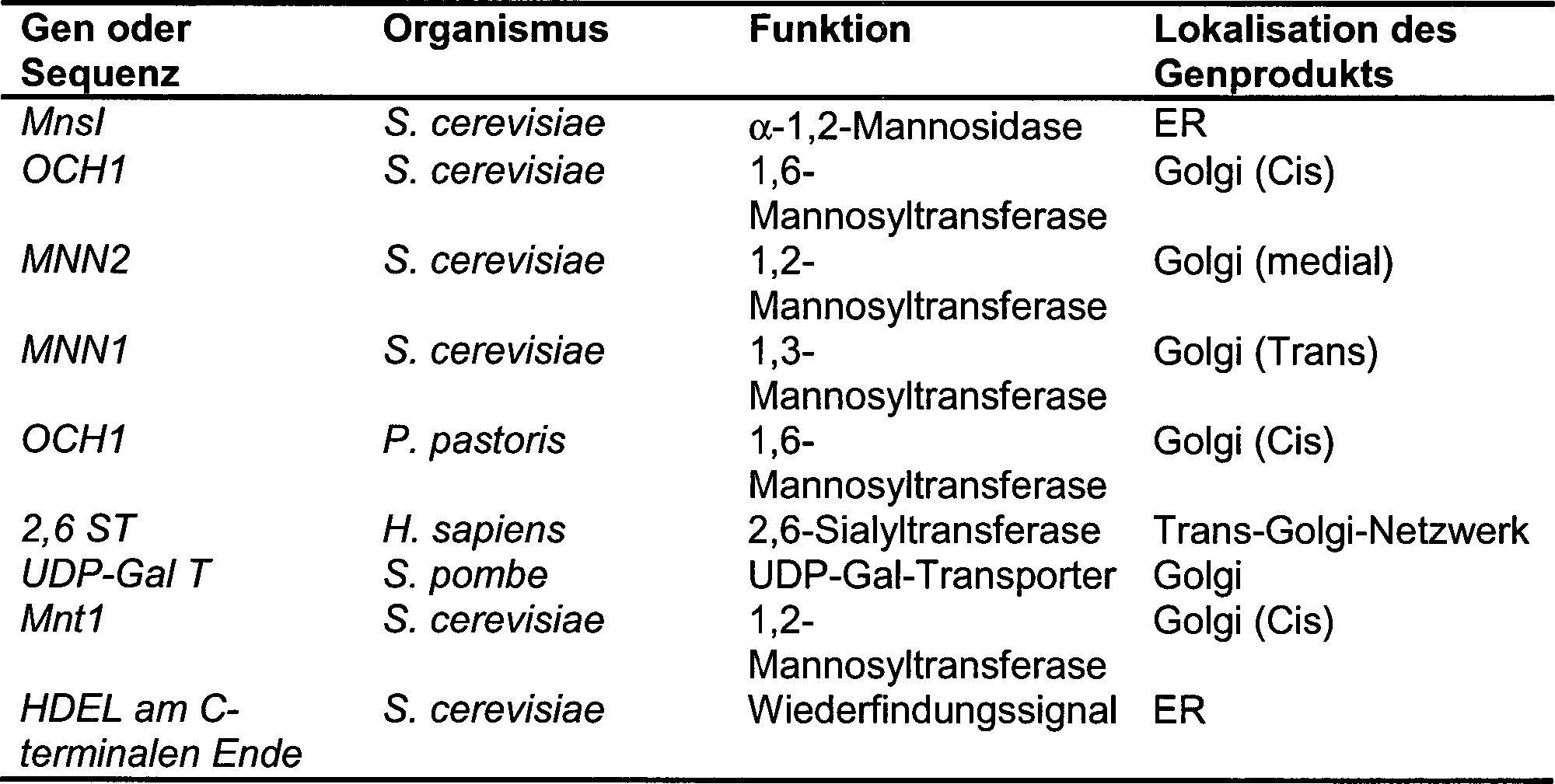
- In jedem Fall wird es stark bevorzugt, dass Signalsequenzen ausgewählt werden, die für die enzymatische(n) Aktivität oder Aktivitäten geeignet sind, die in dem Wirt eingefügt werden sollen. Für die Entwicklung eines modifizierten Mikroorganismus, der in der Lage ist nascente N-Glycane terminal zu sialysieren, ein Prozess, der in Menschen in dem späten Golgi auftritt, ist es zum Beispiel wünschenswert eine Unter-Bibliothek von Signalsequenzen zu verwenden, die von späten Golgi-Proteinen abstammt. In ähnlicher Weise ist das Trimmen von Man8GlcNAc2 durch eine α-1,2-Mannosidase zum Erhalt von Man5GlcNAc2 in Menschen ein früher Schritt bei der komplexen N-Glycan-Bildung. Es ist deshalb wünschenswert, dass man erreicht, dass diese Umsetzung in dem ER oder frühen Golgi eines manipulierten Wirtsorganismus auftritt. Es wird eine Unter-Bibliothek verwendet, die ER- und frühe Golgi-Zurückhaltesignale codiert.
- In einer bevorzugten Ausführungsform wird dann durch die Ligierung einer Unter-Bibliothek, die DNA enthält, die Signalsequenzen codiert, mit einer Unter-Bibliothek, die DNA enthält, die Glycosylierungs-Enzyme oder katalytisch aktive Fragmente davon codiert, im Leserahmen eine DNA-Bibliothek konstruiert. Die resultierende Bibliothek beinhaltet synthetische Gene, die Fusionsproteine codieren. In einigen Fällen ist es wünschenswert an dem N-terminalen Ende eines Fusionsproteins oder in anderen Fällen am C-terminalen Ende eine Signalsequenz bereitzustellen. In einigen Fällen können Signalsequenzen innerhalb des offenen Leserahmens eines Enzyms eingefügt werden, vorausgesetzt die Proteinstruktur individuell gefalteter Domänen wird nicht zerstört.
- Das Verfahren ist am wirkungsvollsten wenn eine DNA-Bibliothek, die in den Wirt transformiert wird, eine große Vielfalt von Sequenzen enthält, wobei die Wahrscheinlichkeit erhöht wird, dass mindestens eine Transformante den gewünschten Phänotyp zeigen wird. Entsprechend kann eine DNA-Bibliothek oder eine bestehende Unter-Bibliothek vor der Transformation einer oder mehreren Runden eines Gen-Shuffling, einer fehlergeneigten PCR oder in vitro-Mutagenese unterzogen werden.
- Transformation
- Die DNA-Bibliothek wird dann in den Wirtsorganismus transformiert. In Hefe kann jedes beliebige geeignete Verfahren des DNA-Transfers verwendet werden, wie Elektroporation, das Lithiumchlorid-Verfahren oder das Sphäroplasten-Verfahren. Um einen stabilen Stamm zu produzieren, der zur Fermentation in hoher Dichte geeignet ist, ist es wünschenswert die DNA-Konstrukte der Bibliothek in das Wirtsgenom zu integrieren. In einer bevorzugten Ausführungsform erfolgt die Integration über homologe Rekombination, wobei Verfahren verwendet werden, die im Stand der Technik bekannt sind. Es werden zum Beispiel DNA-Elemente der Bibliothek mit flankierenden Sequenzen bereitgestellt, die homolog sind zu Sequenzen des Wirtsorganismus. Auf diese Art und Weise findet die Integration an einer bestimmten Stelle in dem Wirtsgenom statt, ohne erwünschte oder notwendige Gene zu zerstören. In einer speziell bevorzugten Ausführungsform wird die DNA der Bibliothek an die Stelle eines unerwünschten Gens in einem Wirtschromosom integriert, was das Zerstören oder die Deletion des Gens bewirkt. Zum Beispiel erlaubt die Integration an die Stellen der OCH1-, MNN1- oder MNN4-Gene die Expression der gewünschten DNA der Bibliothek, während die Expression von Enzymen verhindert wird, die bei der Hefe-Hypermannosylierung von Glycoproteinen beteiligt sind. In anderen Ausführungsformen kann die DNA der Bibliothek mit Hilfe eines Chromosoms, Plasmids, retroviralen Vektors oder durch zufällige Integration in das Wirtsgenom in den Wirt eingebracht werden. In jedem Fall ist es im Allgemeinen wünschenswert mit jedem DNA-Konstrukt der Bibliothek mindestens ein selektionierbares Markergen einzuschließen, das die leichte Selektion von Wirtsorganismen erlaubt, die stabil transformiert worden sind. Wiederverwendbare Marker-Gene wie ura3, auf oder gegen das selektioniert werden kann, sind speziell geeignet.
- Selektionsverfahren
- Nach der Transformation des Wirtsstammes mit der DNA-Bibliothek werden Transformanten ausgewählt, die den gewünschten Glycosylierungs-Phänotyp zeigen. Die Selektion kann in einem einzelnen Schritt oder durch eine Reihe phänotypischer Anreicherungs- und/oder Verarmungs-Schritte durchgeführt werden, wobei jeder beliebige aus einer Vielzahl von Tests oder Nachweisverfahren verwendet werden kann. Die phänotypische Charakterisierung kann von Hand durchgeführt werden oder einen automatischen Hochdurchsatz-Durchmusterungs-Apparat verwenden. Im Allgemeinen zeigt ein Wirtsorganismus Protein-N-Glycane auf der Zelloberfläche, wo verschiedene Glycoproteine lokalisiert sind. Entsprechend können intakte Zellen nach einem gewünschten Glycosylierungs-Phänotyp durchmustert werden, indem die Zellen einem Lektin oder Antikörper ausgesetzt werden, der spezifisch an das gewünschte N-Glycan bindet. Eine große Bandbreite Oligosaccharid-spezifischer Lektine ist kommerziell erhältlich (EY Laboratories, San Mateo, CA). Alternativ dazu sind Antikörper gegen spezifische menschliche oder tierische N-Glycane kommerziell erhältlich oder können unter Verwendung von Standard-Verfahren hergestellt werden. Ein geeignetes Lektin oder ein geeignerter Antikörper kann mit einem Reporter-Molekül wie einem Chromophor, Fluorophor, einem Radioisotop oder einem Enzym, das ein chromogenes Substrat hat, konjugiert sein (Guillen et al., 1998. Proc. Natl. Acad. Sci. USA 95 (14): 7888–7892). Die Durchmusterung kann dann durchgeführt werden, indem analytische Verfahren wie Spektrophotometrie, Fluorimetrie, Fluoresenz-aktivierte Zell-Sortierung oder Scintillationszählung verwendet werden. In anderen Fällen kann es notwendig sein isolierte Glycoproteine oder N-Glycane von transformierten Zellen zu analysieren. Die Protein-Isolierung kann unter Verwendung von im Stand der Technik bekannten Verfahren durchgeführt werden. In den Fällen, in denen ein isoliertes N-Glycan erforderlich ist, kann ein Enzym wie eine Endo-β-N-Acetyl-Glucosaminidase (Genzyme Co., Boston, MA) verwendet werden, um die N-Glycane von Glycoproteinen zu spalten. Isolierte Proteine oder N-Glycane können dann durch Flüssigkeits-Chromatographie (z.B. HPLC), Massen-Spektroskopie oder andere geeignete Verfahren analysiert werden. Das US-Patent Nr. 5,595,900 lehrt einige Verfahren, mit denen Zellen mit gewünschten extrazellulären Kohlenhydrat-Strukturen identifiziert werden können. Vor der Auswahl einer gewünschten Transformante kann es wünschenswert sein, die transformierte Population um diejenigen Zellen zu verarmen, die unerwünschte Phänotypen besitzen. Wenn das Verfahren zum Beispiel verwendet wird, um eine funktionelle Mannosidase-Aktivität in Zellen einzubringen, wird die gewünschte Transformante niedrigere Spiegel an Mannose im zellulärem Glycoprotein besitzen. Exponiert man die transformierte Population mit einem letalen Radioisotop von Mannose im Medium, verarmt man die Population um Transformanten, die den ungewünschten Phänotyp besitzen, d.h. hohe Spiegel an eingebauter Mannose. Alternativ dazu kann ein cytotoxisches Lektin oder ein cytotoxischer Antikörper verwendet werden, das/der gegen ein unerwünschtes N-Glycan gerichtet ist, um eine transformierte Population um unerwünschte Phänotypen zu verarmen.
- Verfahren zur Bereitstellung von Zucker-Nucleotid-Vorläufern an den Golgi-Apparat
- Für eine Glycosyltransferase, die in dem Golgi zufriedenstellend funktionieren soll, ist es notwendig, dass dem Enzym eine ausreichende Konzentration eines geeigneten Nucleotid-Zuckers bereitgestellt wird, der den Hoch-Energie-Spender der Zuckereinheit darstellt, die an ein nascierendes Glycoprotein angefügt wird. Diese Nucleotid-Zucker werden dem geeigneten Kompartiment bereitgestellt, indem ein exogenes Gen, das einen Zucker-Nucleotid-Transporter codiert, in dem Wirts-Mikroorganismus exprimiert wird. Die Wahl des Transporter-Enzyms wird durch die Natur der verwendeten exogenen Glycosyltransferase beeinflusst. Eine GlcNAc-Transferase zum Beispiel kann einen UDP-GlcNAc-Transporter benötigen, eine Fucosyltransferase kann einen GDP-Fucose-Transporter benötigen, eine Galactosyltransferase kann einen UDP-Galactose-Transporter benötigen oder eine Sialyltransferase kann einen CMP-Sialinsäure-Transporter benötigen.
- Das zugegebene Transporter-Protein befördert einen Nucleotid-Zucker vom Cytosol in den Golgi-Apparat, wo der Nucleotid-Zucker von der Glycosyltransferase umgesetzt wird, z.B. um ein N-Glycan zu verlängern. Die Umsetzung setzt ein Nucleosid-Diphosphat oder -Monophosphat frei, z.B. UDP, GDP oder CMP. Da die Ansammlung eines Nucleosid-Diphosphats die weitere Aktivität einer Glycosyltransferase hemmt, ist es häufig auch wünschenswert eine exprimierte Kopie eines Gens bereitzustellen, das eine Nucleotid-Diphosphatase codiert. Die Diphosphatase (spezifisch für UDP oder GDP, soweit angemessen) hydrolysiert das Diphosphonucleosid, wobei ein Nucleosid-Monophosphat und anorganisches Phosphat erhalten werden. Das Nucleosid-Monophosphat hemmt die Glycotransferase nicht und wird in jedem Fall durch ein endogenes zelluläres System aus dem Golgi exportiert. Geeignete Transporter-Enzyme, die typischerweise aus einem Säuger stammen, sind nachfolgend beschrieben.
- Beispiele
- Die Verwendung des vorstehenden allgemeinen Verfahrens kann durch Bezugnahme auf die nachfolgenden nicht einschränkenden Beispiele verstanden werden. Beispiele von bevorzugten Ausführungsformen sind auch in Tabelle 6 zusammengefasst.
- Beispiel 1: Manipulation von P. pastoris mit α-1,2-Mannosidase, um Interferon herzustellen.
- Eine α-1,2-Mannosidase ist erforderlich, um Man8GlcNAc2 zu trimmen, wobei Man5GlcNAc2 erhalten wird, ein notwendiges Zwischenprodukt für die Bildung eines komplexen N-Glycans. Eine OCH1-Mutante von P. pastoris wird manipuliert, um menschliches Interferon-β unter der Kontrolle eines aox-Promotors zu exprimieren. Durch Ligierung der katalytischen Domäne von menschlicher Mannosidase IB (eine α-1,2-Mannosidase) im Leserahmen mit einer Unter-Bibliothek, die Sequenzen beinhaltet, die frühe Golgi-Lokalisierungs-Peptide codieren, wird eine DNA-Bibliothek konstruiert. Die DNA-Bibliothek wird dann in den Wirtsorganismus transformiert, wobei eine genetisch gemischte Population resultiert, in der einzelne Transformanten sowohl Interferon-β als auch ein synthetisches Mannosidase-Gen aus der Bibliothek exprimieren. Einzelne Transformanten-Kolonien werden gezüchtet und die Herstellung von Interferon wird durch die Zugabe von Methanol induziert. Unter diesen Bedingungen beinhalten mehr als 90% des sekretierten Proteins Interferon-β. Die Überstande werden durch C18-Silica-Umkehrphasen-Chromatographie gereinigt, um Salze und niedermolekulare Fremdstoffe zu entfernen. Gewünschte Transformanten, die angemessen zielgerichtete aktive α-1,2-Mannosidase exprimieren, stellen Interferon-β einschließlich N-Glycanen der Struktur Man5GlcNAc2 her, die verglichen mit dem Interferon des parentalen Stammes eine reduzierte molekulare Masse besitzen. Die gereinigten Überstande, einschließlich Interferon-β, werden durch MALDI-TOF- Massenspektroskopie analysiert und Kolonien werden identifiziert, die die gewünschte Form von Interferon-β exprimieren.
- Beispiel 2: Manipulation eines Stammes zur Expression von GlcNAc-Transferase I
- GlcNAc-Transferase I-Aktivität ist für die Reifung von komplexen N-Glycanen erforderlich. Man5GlcNAc2 kann ausschließlich durch Mannosidase II getrimmt werden, ein notwendiger Schritt bei der Bildung menschlicher Glycoformen nach dem Anhängen von GlcNAc an den terminalen α-1,3-Mannose-Rest durch GlcNAc-Transferase I (Schachter, 1991 Glycobiology 1(5): 453–461). Entsprechend wird eine Bibliothek hergestellt, die DNA-Fragmente enthält, die geeignete zielgerichtete GlcNAc-Transferase 1-Gene codieren. Der Wirtsorganismus ist ein Stamm, z.B. eine Hefe, der für Hypermannosylierung defizient ist (z.B. eine OCH1-Mutante), das Substrat UDP-GlcNAc in dem Golgi und/oder dem ER bereitstellt und N-Glycane der Struktur Man5GlcNAc2 in dem Golgi und/oder dem ER bereitstellt. Nach Transformation des Wirts mit der DNA-Bibliothek werden die Transformanten nach denjenigen durchmustert, die die höchste Konzentration an terminalem GlcNAc auf der Zell-Oberfläche besitzen oder alternativ dazu das Protein sekretieren, das den höchsten terminalen GLcNAc-Gehalt besitzt. Eine solche Durchmusterung wird unter Verwendung eines visuellen Verfahrens (z.B. eine Färbemethode), eines spezifischen terminalen GLcNAc-bindenden Antikörpers oder eines Lektins durchgeführt. Alternativ dazu zeigen die gewünschten Transformanten eine reduzierte Bindung bestimmter Lektine, die für terminale Mannose-Reste spezifisch sind.
- Beispiel 3: Manipulation von Stämmen mit einer Mannosidase II
- In einem anderen Beispiel ist es erwünscht die zwei verbleibenden terminalen Mannosen aus der Struktur GlcNAcMan5GlcNAc2 durch Wirkung einer Mannosidase II zu entfernen, um eine menschliche Glycoform in einem Mikroorganismus zu erzeugen. Eine DNA-Bibliothek, die Sequenzen beinhaltet, die Cis- und mediale Golgi-Lokalisations-Signale codieren, wird im Leserahmen mit einer Bibliothek fusioniert, die katalytische Domänen von Mannosidase II codiert. Der Wirtsorganismus ist ein Stamm, z.B. eine Hefe, der für Hypermannosylierung defizient ist (z.B. eine OCH1-Mutante) und N-Glycane bereitstellt, die die Struktur GlcNAcMan5GlcNAc2 in dem Golgi und/oder dem ER besitzen. Nach Transformation werden Organismen selektioniert, die den gewünschten Glycosylierungs-Phänotyp besitzen. In einem Verfahren wird ein in vitro-Test verwendet. Die gewünschte Struktur GlcNAcMan3GlcNAc2 (jedoch nicht unerwünschtes GlcNAcMan5GlcNAc2) ist ein Substrat für das Enzym GlcNAc-Transferase II. Entsprechend können einzelne Kolonien unter Verwendung dieses Enzyms in vitro in Anwesenheit des Substrats, UDP-GlcNAc, getestet werden. Die Freisetzung von UDP wird entweder durch HPLC oder einen enzymatischen Test für UDP bestimmt. In einer alternativen Ausführungsform wird radioaktiv markiertes UDP-GlcNAc verwendet.
- Die vorangehenden in vitro-Tests werden in geeigneter Weise mit einzelnen Kolonien durchgeführt, wobei eine Hochdurchsatz-Durchmusterungs-Apparatur verwendet wird. In einer alternativen Ausführungsform wird ein Lektin-Bindungstest verwendet. In diesem Fall erlaubt die reduzierte Bindung von Lektinen, die für terminale Mannosen spezifisch sind, die Selektion von Transformanten, die den gewünschten Phänotyp besitzen. Galantus nivalis-Lektin zum Beispiel bindet spezifisch an terminale α-1,3-Mannose, deren Konzentration in Anwesenheit von funktionell exprimierter Mannosidase II-Aktivität reduziert ist. In einem geeigneten Verfahren wird G. nivalis-Lektin verwendet, das an einen festen Agarose-Träger (erhältlich von Sigma Chemical, St. Louis, MO) angeheftet ist, um die transformierte Population an Zellen zu verarmen, die hohe Spiegel an terminaler α-1,3-Mannose aufweisen.
- Beispiel 4: Manipulation von Organismen, die Sialyltransferase exprimieren
- Die Enzyme α-2,3-Sialyltransferase und α-2,6-Sialyltransferase fügen terminale Sialinsäure an Galactose-Reste in nascierenden menschlichen N-Glycanen an, was zu reifen Glycoproteinen führt. Im Menschen treten die Reaktionen in dem Trans-Golgi oder dem TGN auf. Entsprechend wird durch die Fusion von Sequenzen, die katalytische Domänen von Sialyltransferase codieren, im Leserahmen mit Sequenzen, die Trans-Golgi- oder TGN-Lokalisierungs-Signale codieren, eine DNA-Bibliothek erzeugt. Der Wirtsorganismus ist ein Stamm, z.B. eine Hefe, der für Hypermannosylierung defizient ist (z.B. OCH1-Mutante), die N-Glycane bereitstellt, die terminate Galactose-Reste in dem Trans-Golgi oder dem TGN besitzen, und eine ausreichende Konzentration an CMP-Sialinsäure in dem Trans-Golgi oder TGN bereitstellt. Nach der Transformation werden Transformanten selektioniert, die den gewünschten Phänotyp besitzen, wobei ein fluoreszierender Antikörper verwendet wird, der für N-Glycane spezifisch ist, die eine terminale Sialinsäure besitzen.
- Beispiel 5: Verfahren zur Manipulation von Stämmen, um einen UDP-GlcNAc-Transporter zu exprimieren
- Die cDNA von menschlichem Golgi-UDP-GlcNAc-Transporter wurde von Ishida und Mitarbeitern cloniert (Ishida; N. et al. 1999 J. Biochem. 126 (1): 68–77). Guillen und Mitarbeiter haben den Hunde-Nieren-Golgi-UDP-GlcNAc-Transporter durch phänotypische Korrektur einer Kluyveromyces lactis-Mutante cloniert, die für den Golgi-UDP-GlcNAc-Transport defizient war (Guillen, E. et al. 1998). So besitzt ein Säuger-Golgi-UDP-GlcNAc-Transporter-Gen alle notwendigen Informationen dass das Protein funktionell exprimiert und zu den Golgi-Apparat von Hefe gelenkt wird.
- Beispiel 6: Verfahren zur Manipulation von Stämmen, um einen GDP-Fucose-Transporter zu exprimieren
- Der Rattenleber-Golgi-Membran-GDP-Fucose-Transporter wurde von Puglielle, L. und C. B. Hirschberg 1999 J. Biol. Chem. 274 (59): 35596–35600 identifiziert und aufgereinigt. Das entsprechende Gen kann unter Verwendung von Standard-Verfahren wie N-terminale Sequenzierung und Southern-Blottverfahren unter Verwendung einer degenerierten DNA-Probe identifiziert werden. Das intakte Gen wird dann in einem Wirtsorganismus exprimiert, der ebenfalls eine Fucosyltransferase exprimiert.
- Beispiel 7: Verfahren zur Manipulation von Stämmen, um einen UDP-Galactose-Transporter zu exprimieren
- Ein menschlicher UDP-Galactose (UDP-Gal)-Transporter wurde cloniert, und es wurde gezeigt, dass dieser in S. cerevisiae aktiv ist (Kainuma, M., et al. 1999 Glycobiology 9 (2): 133–141). Ein zweiter menschlicher UDP-Galactose-Transporter (hUGT1) wurde cloniert und in Ovarzellen des Chinesischen Hamsters funktionell exprimiert. Aoki, K., et al. 1999 J. Biochem. 126 (5): 940–950. Auf ähnliche Weise haben Segawa und Mitarbeiter einen UDP-Galactose-Transporter von Schizosaccharomyces pombe cloniert (Segawa, H., et al. 1999 Febs Letters 451 (3): 295–298).
- CMP-Sialinsäure-Transporter
- Ein menschlicher CMP-Sialinsäure-Transporter (hCST) wurde von Aoki und Mitarbeitern (1999) cloniert und in Lec 8 CHO-Zellen exprimiert. Die molekulare Clonierung des Hamster-CMP-Sialinsäure-Transporters wurde ebenfalls erreicht (Eckhardt and Gerardy Schahn 1997 Eur. J. Biochem. 248 (1): 187–192). Die funktionelle Expression des murinen CMP-Sialinsäure-Transporters wurde von Berninsone, P., et al. 1997 J. Biol. Chem. 272 (19): 12616–12619 in Saccharomyces cerevisiae erreicht.
- Tabelle 6. Beispiele für bevorzugte Ausführungsformen des Verfahrens zur Modifikation von Glycosylierung in einem eukaryontischen Mikroorganismus, z.B. Pichia pastoris

-

- Tabelle 7: Quellen für DNA- und Protein-Sequenzen
-
- 1. European Bioinformatics Institute (EBI) ist ein Zentrum für Forschung und Dienstleistungen in der Bioinformatik: http://www.ebi.ac.uk/
- 2. Swissport Datenbank: http://www.expasy.ch/spr
- 3. Liste bekannter Glycosyltransferasen und deren Ursprung.
- β1,2-(GnT I) EC 2.4.1.101
-
- 4. Menschliche cDNA, Kumar et al. (1990) Proc. Natl. Acad. Sci. USA 87: 9948–9952
- 5. Menschliches Gen, Hull et al. (1991) Biochem. Biophys. Res. Commun. 176: 608–615
- 6. Maus-cDNA, Kumar et al. (1992) Glycobiology 2: 383–393
- 7. Maus-Gen, Pownall et al. (1992) Genomics 12: 699–704
- 8. Maus-Gen (5'-flankierend, nicht codierend), Yang et al. (1994) Glycobiology 5: 703–712
- 9. Kaninchen-cDNA, Sarkar et al. (1991) Proc. Natl. Acad. Sci. USA 88: 234–238
- 10. Ratten-cDNA, Fukuda et al. (1994) Biosci. Biotechnol. Biochem. 58: 200–201
- 1,2-(GnT II) EC 2.4.1.143
-
- 11. Menschliches Gen, Tan et al. (1995) Eur. J. Biochem. 231: 317–328
- 12. Ratten-cDNA, D'Agostaro et al. (1995) J. Biol. Chem. 270: 15211–15221
- 13. β1,4 (GnT III), EC 2.4.1.144
- 14. Menschliche cDNA, Ihara et al. (1993) J. Biochem. 113: 692–698
- 15. murines Gen, Bhaumik et al. (1995) Gene 164: 295–300
- 16. Ratten-cDNA, Nishikawa et al. (1992) J. Biol. Chem. 267: 18199–18204
- β1,4-(GnT IV) EC 2.4.1.145
-
- 17. Menschliche cDNA, Yoshida et al. (1998) Glycoconjugate Journal 15: 1115–1123
- 18. Rinder-cDNA, Minowa et al., Europäisches Patent
EP 0 905 232 (31,6 (GnT V) EC 2.4.1.155 - 19. Menschliche cDNA, Saito et al. (1994) Biochem. Biophys. Res. Commun. 198: 318–327
- 20. Ratten-cDNA, Shoreibah et al. (1993) J. Biol. Chem. 268: 15381–15385
- β1,4-Galactosyltransferase, EC 2.4.1.90 (LacNAc-Synthetase) EC
- 2.4.1.22 (Lactose-Synthetase)
-
- 21. Rinder-cDNA, D'Agostaro et al. (1989) Eur. J. Biochem. 183: 211–217
- 22. Rinder-cDNA (partiell), Narimatsu et al. (1986) Proc. Natl. Acad. Sci. USA 83: 4720–4724
- 23. Rinder-cDNA (partiell), Masibay & Qasba (1989) Proc. Natl. Acad. Sci. USA 86: 5733–5377
- 24. Rinder-cDNA (5'-Ende), Russo et al. (1990) J. Biol. Chem. 265: 3324
- 25. Huhn-cDNA (partiell), Ghosh et al. (1992) Biochem. Biophys. Res. Commun. 1215–1222
- 26. Menschliche cDNA, Masri et al. (1988) Biochem. Biophys. Res. Commun. 157: 657–663
- 27. Menschliche cDNA, (HeLa-Zellen). Watzele & Berger (1990) Nucl. Acids Res. 18: 7174
- 28. Menschliche cDNA, (partiell) Uejima et al (1992) Cancer Res. 52: 6158–6163
- 29. Menschliche cDNA, (Karzinom) Appert et al. (1986) Biochem. Biophys. Res. Commun. 139: 163–168
- 30. Menschliches Gen, Mengle-Gaw et al. (1991) Biochem. Biophys. Res. Commun. 176: 1269–1276
- 31. murine cDNA, Nakazawa et al. (1988) J. Biochem. 104: 165–168
- 32. murine cDNA, Shaper et al. (1988) J. Biol. Chem. 263: 10420–10428
- 33. murine cDNA (neu), Uehara & Muramatsu, nicht veröffentlicht
- 34. murines Gen, Hollis et al. (1989) Biochem. Biophys. Res. Commun. 162: 1069–1075
- 35. Rattenprotein (partiell), Bendiak et al. (1993) Eur. J. Biochem. 216: 405–417
- 2,3-Sialyltransferase, (ST3Gal II) (N-gekoppelt) (Gal-1,3/4-GlcNAc) EC 2.4.99.6
-
- 36. Menschliche cDNA, Kitagawa & Paulson (1993) Biochem. Biophys. Res. Commun. 194: 375–382
- 37. Ratten-cDNA, Wen et al. (1992) J. Biol. Chem. 267: 21011–21019
- 2,6-Sialyltransferase, (ST6Gal I) EC 2.4.99.1
-
- 38. Huhn, Kurosawa et al. (1994) Eur. J. Biochem 219: 375–381
- 39. Menschliche cDNA (partiell), Lance et al. (1989) Biochem. Biophys. Res. Commun. 164: 225–232
- 40. Menschliche cDNA, Grundmann et al. (1990) Nucl. Acids Res. 18: 667
- 41. Menschliche cDNA, Zettlmeisl et al. (1992) Patent EPO475354-A/3
- 42. Menschliche cDNA, Stamenkovic et al. (1990) J. Exp. Med. 172: 641–643 (CD75)
- 43. Menschliche cDNA, Bast et al. (1992) J. Cell Biol. 116: 423–435
- 44. Menschliches Gen (partiell), Wang et al. (1993) J. Biol. Chem. 268: 4355–4361
- 45. Menschliches Gen (5'-flankierend), Aasheim et al. (1993) Eur. J. Biochem. 213: 467–475
- 46. Menschliches Gen (Promotor), Aas-Eng et al. (1995) Biochim. Biophys. Acta 1261: 166–169
- 47. Maus-cDNA, Hamamoto et al. (1993) Bioorg. Med. Chem. 1: 141–145
- 48. Ratten-cDNA, Weinstein et al. (1987) J. Biol. Chem. 262: 17735–17743
- 49. Ratten-cDNA (Transkript-Fragmente), Wang et al. (1991) Glycobiology 1: 25–31, Wang et al. (1990) J. Biol. Chem. 265: 17849–17853
- 50. Ratten-cDNA (5'-Ende), O'Hanlon et al. (1989) J. Biol. Chem. 264: 17389–17394; Wang et al. (1991) Glycobiology 1: 25–31
- 51. Ratten-Gen (Promotor), Svensson et al. (1990) J. Biol. Chem. 265: 20863–20688
- 52. Ratten-mRNA (Fragmente), Wen et al. (1992) J. Biol. Chem. 267: 2512–2518
- Zusätzliche Verfahren und Reagenzien, die in den Verfahren zur Modifikation der Glycolysierung verwendet werden können, sind in der Literatur beschrieben, wie das US-Patent Nr. 5,955,422, US-Patent Nr. 4,775,622, US-Patent Nr. 6,017,743, US-Patent Nr. 4,925,796, US-Patent Nr. 5,766,910, US-Patent Nr. 5,834,251, US-Patent Nr. 5,910,570, US-Patent Nr. 5,849,904, US-Patent Nr. 5,955,347, US-Patent Nr. 5,962,294, US-Patent Nr. 5,135,854, US-Patent Nr. 4,935,349, US-Patent Nr. 5,707,828 und US-Patent Nr. 5,047,335.
- Geeignete Hefe-Expressions-Systeme können von Quellen wie die American Type Culture Collection, Rockville, MD erhalten werden. Vektoren sind von einer Vielzahl von Quellen kommerziell erhältlich.
Claims (34)
- Wirtszelle, die ein unizellulärer oder filamentöser Pilz ist, und keine alpha-1,6-Mannosyltransferaseaktivität, bezogen auf das N-Glycan auf einem Glycoprotein, aufweist, und die in ihrem Endoplasmatischen Reticulum (ER) oder Golgi-Apparat ein Hybridenzym hat, das ausgewählt ist, um optimale Aktivität im ER oder Golgi der Wirstzelle zu haben, so dass die Wirtszelle 50 bis 100 Mol% Man5GlcNAc2 auf einem Substrat-Glycoprotein bilden kann, wobei das Hybridenzym umfasst: (a) eine exogene Mannosidase-katalytische Domäne mit optimaler Aktivität im ER oder Golgi bei einem pH zwischen 5,1 und 8,0; verbunden mit (b) einem zellulären Zielsignal-Peptid, das normalerweise nicht mit der katalytischen Domäne von (a) verbunden ist, wobei das zelluläre Zielsignal-Peptid die exogene Mannosidase-katalytische Domäne in das ER oder den Golgi-Apparat bringt.
- Wirtszelle nach Anspruch 1, wobei die katalytische Domäne α-1,2-Mannosidaseaktivität hat.
- Wirtszelle nach Anspruch 1 oder 2, wobei die Mannosidase eine α-1,2-Mannosidase-katalytische Domäne umfasst, die von einer Maus, einem Menschen, Lepidoptera, Aspergillus nidulans, Xanthomonas manihotis oder Bacillus sp. stammt.
- Wirtszelle nach einem der Ansprüche 1 bis 3, wobei die Mannosidase optimale Aktivität bei einem pH zwischen 5,9 und 7,5 hat.
- Wirtszelle nach einem der Ansprüche 1 bis 4, die weiterhin ein oder mehrere exogenes) Enzyme) exprimiert/exprimieren, ausgewählt aus der Gruppe bestehend aus Mannosidasen, Glycolsyltransferasen und Glycosidasen.
- Wirtszelle nach Anspruch 5, wobei mindestens ein zusätzliches exogenes Enzym für die Herstellung eines Glycoproteins lokalisiert wird, indem ein Fusionsprotein zwischen einer katalytischen Domäne des exogenen Enzyms und einem zellulären Zielsignal-Peptid gebildet wird, das normalerweise nicht mit der katalytischen Domäne verbunden ist, die es in das Endoplasmatische Reticulum, den frühen, mittleren oder späten Golgi oder in das trans-Golgi Netzwerk der Wirttzelle bringt.
- Wirtszelle nach Anspruch 6, wobei die katalytische Domäne von einer Glycosidase oder Glycosyltransferase stammt, ausgewählt aus GnT I, GnT II, GnT III, GnT IV, GnT V, GnT VI.
- Wirtszelle nach einem der Ansprüche 5 bis 7, die zusätzlich ein oder mehrere exogenes) Enzyme) exprimiert/exprimieren, ausgewählt aus der Gruppe bestehend aus UDP-GlcNAc-Transferase, UDP-GlcNAc-Transporter und Nucleotid-Diphosphatasen.
- Wirtszelle nach einem der Ansprüche 5 bis 8, die zusätzlich ein oder mehrere exogenes) Enzyme) exprimiert/exprimieren, ausgewählt aus: einer UDP-spezifischen Diphosphatase und einer GDP-spezifischen Diphosphatase.
- Wirtszelle nach einem der Ansprüche 1 bis 9, erhalten durch Einbringen von Nucleisäuremolekülen, welche die Enzyme codieren.
- Wirtszelle nach Anspruch 10, wobei mindestens eines der Nucleinsäuremoleküle, welche die Enzyme codieren, in das Wirtszell-Chromosom integriert ist.
- Wirtszelle nach einem der Ansprüche 1 bis 11, die ausgewählt ist aus der Gruppe bestehend aus Pichia pastoris, Pichia finlandica, Pichia trehalophila, Pichia koclamae, Pichia membranaefaciens, Pichia opuntiae, Pichia thermotolerans, Pichia salictaria, Pichia guercuum, Pichia pijperi, Pichia stiptis, Pichia methanolica, Pichia sp., Saccharomyces cerevisae, Saccharomyces sp., Hansenula polymorpha, Kluyveromyces sp., Candida Albicans, Aspergillus nidulans und Trichoderma reesei.
- Wirtszelle nach einem der Ansprüche 1 bis 12, die zusätzlich keine Aktivität eines oder mehrerer Enzyms/Enzyme hat, ausgewählt aus der Gruppe bestehend aus Mannosyltransferasen und Phosphomannosyltransferasen.
- Wirtszelle nach Anspruch 13, die zusätzlich kein Enzym exprimiert, ausgewählt aus der Gruppe bestehend aus 1,6-Mannosyltransferase; 1,3-Mannosyltransferase; und 1,2-Mannosyltransferase bezogen auf das N-Glycan.
- Wirtszelle nach einem der Ansprüche 1 bis 14, wobei der Wirt eine OCH1-Mutante von P. pastoris ist.
- Verfahren zur Herstellung einer Wirtszelle nach einem der Ansprüche 1 bis 15, wobei das Verfahren den Schritt des Transformierens des unizellulären oder filamentösen Pilzes mit einem Nucleinsäuremolekül umfasst, das ein Hybridenzym codiert, das ausgewählt ist, um optimale Aktivität im ER oder Golgi der Wirtszelle zu haben, so dass die Wirtszelle 50 bis 100 Mol% Man5GlcNAc2 auf einem Substrat-Glycoprotein bilden kann, wobei das Hybridenzym umfasst: (a) eine exogene Mannosidase-katalytische Domäne mit optimaler Aktivität im ER oder Golgi bei einem pH zwischen 5,1 und 8,0; verbunden mit (b) einem zellulären Zielsignal-Peptid, dass normalerweise nicht mit der katalytischen Domäne von (a) verbunden ist, wobei das zelluläre Zielsignal-Peptid die exogene Mannosidase-katalytische Domäne in das ER oder den Golgi-Apparat bringt.
- Verfahren zur Herstellung eines Glycoproteins, umfassend Exprimieren des Glycoproteins in einer Wirtszelle nach einem der Ansprüche 1 bis 15.
- Verfahren nach Anspruch 17, wobei das Glycoprotein N-Glycane mit weniger als sechs Mannoseresten umfasst.
- Verfahren nach Anspruch 17 oder 18, wobei das Glycoprotein N- Acetylglucosamin umfasst.
- Verfahren nach einem der Ansprüche 17 bis 19, wobei das Glycoprotein mindestens eine Oligosaccharid-Verzweigung umfasst, welche die Struktur GlcNAc-Man umfasst.
- Verfahren nach einem der Ansprüche 17 bis 20, wobei das Glycoprotein ausgewählt ist aus der Gruppe bestehend aus Erythropoietin, Interferon-α, Interferon-β, Interferon-γ, Interferon-ω, Granulocyten-CSF, Faktor VIII, Faktor IX, menschliches Protein C, löslicher IgE-Rezeptor-α-Kette, IgG, IgM, Urokinase, Chymase, Harnstoff-Trypsin-Inhibitor, IGF-bindendem Protein, epidermalem Wachstumsfaktor, Wachstumshormon-freisetzendem Faktor, Annexin V-Fusionsprotein, Angiostatin, vaskulärem endothelialem Wachstumsfaktor-2, myeloidem Vorläufer-Inhibitor-Faktor-1 und Osteoprotegerin.
- Verfahren nach einem der Ansprüche 17 bis 21, weiterhin umfassend den Schritt des Erntens des Glycoproteins vom Wirt.
- Verfahren nach Anspruch 22, weiterhin umfassend den Schritt das geerntete Glycoprotein mindestens einer Glycosilierungsreaktion in vitro zu unterziehen, im Anschluss an das Ernten vom Wirt.
- Verfahren zur Herstellung eines Glycoproteins in einer Wirtszelle, die ein unizellulärer oder filamentöser Pilz ist, und die keine alpha-1,6-Mannosyltransferaseaktivität im Bezug auf das N-Glycan auf einem Glycoprotein aufweist, wobei das Verfahren die Schritte umfasst: (a) Exprimieren des Glycoproteins in einer Population von Wirtszellen, die mit einer DNA-Bibliothek transformiert sind, die mindestens zwei verschiedene genetische Konstrukte umfasst, wobei beide ein DNA-Fragmentumfassen, das ein Hybridenzym codiert, umfassend: (i) eine Glycosilierungsenzym-katalytische Domäne mit optimaler Aktivität im ER oder Golgi bei einem pH zwischen 5,1 und 8,0; verbunden mit (ii) einem zellulären Zielsignal-Peptid, das normalerweise nicht mit der katalytischen Domäne von (i) verbunden ist, wobei das zelluläre Zielsignal-Peptid auf das ER oder den Golgi-Apparat der Wirttzelle abzielt; und (b) Auswählen der Wirtszelle, die ein Glycoprotein mit einem gewünschten Glycosilierungsmuster auf dem Glycoprotein zeigt.
- Verfahren nach Anspruch 24, wobei die Bibliothek mindestens ein Gen oder genetisches Konstrukt umfasst, das vorher einer Technik unterzogen wurde, ausgewählt aus: Gene-Shuffling, in vitro-Mutagenese und fehleranfälliger Polymerase-Ketten-Reaktion (error-prone polymerase chain reaction).
- Verfahren nach Anspruch 24 oder 25, wobei das DNA-Fragment, das die Glycosilierungsenzym-katalytische Domäne codiert, eine Aktivität hat, ausgewählt aus der Gruppe bestehend aus Mannosidase, Glycosidase, UDP-GlcNAc-Transferase, Glycosyltransferase, ausgewählt aus GnT I, GnT II, GnT III, GnT IV, GnT V, GnT VI; und wobei das zelluläre Zielsignal-Peptid das Enzym vorwiegend in einem Wirtszell-Organell lokalisiert, ausgewählt aus der Gruppe bestehend aus endoplasmatischem Reticulum, cis-Golgi, medium-Golgi, und trans-Golgi.
- Verfahren nach Anspruch 24 oder 25, wobei die codierte Glycosylierungsenzym-katalytische Domäne Mannosidaseaktivität hat.
- Verfahren nach Anspruch 27, wobei die Wirtszelle weiterhin GnT I exprimiert.
- Verfahren nach einem der Ansprüche 24 bis 28, wobei die Bibliothek mindestens ein DNA-Fragment umfasst, das ein Wildtyp-Glycosylierungsenzym codiert.
- Verfahren nach einem der Ansprüche 24 bis 29, wobei das Auswählen den Schritt des Untersuchens des glycosilierten Proteins oder isolierten N-Glycans anhand eines oder mehrerer Verfahren umfasst, ausgewählt aus der Gruppe bestehend aus: (a) Massenspektroskopie, wie z.B. MALDI-TOF-MS; (b) Flüssigkeitschromatographie; (c) Charakterisieren von Zellen durch Verwenden eines Fluoreszenz-aktivierten Zell-Sortierers, eines Spektrophotometers, Fluorimeters oder Szintialltionszählers; (d) In-Kontakt-bringen der Wirtszellen mit einem Lektin oder Antikörper mit einer spezifischen Affinität für einen gewünschten Oligosaccharid-Rest; und (e) In-Kontakt-bringen der Zellen mit einem cytotoxischen oder radioaktiven Molekül, ausgewählt aus der Gruppe bestehend aus Zuckern, Antikörpern und Lektinen.
- Transformierte Wirtszelle, hergestellt in dem Verfahren nach einem der Ansprüche 16 oder 24 bis 30.
- Glycoprotein-Zusammensetzung, erhältlich nach einem der Verfahren der Ansprüche 17 bis 30, umfassend 50 bis 100 Mol% von Man5GlcNAc2, in vivo durch GnT I zu GlcNacMan5GlcNAc2 umgesetzt, wobei dem Glycoprotein Galactose, Fucose und terminale Sialinsäure fehlen.
- DNA-Bibliothek, umfassend genetische Konstrukte, wobei die Konstrukte ein DNA-Fragment umfassen, das ein zelluläres Zielsignal-Peptid codiert, das in das Endoplasmatische Reticulum oder den Golgi-Apparat abzielt, und das im Leserahmen mit einem DNA-Fragment verbunden ist, das ein Glycosylierungsenzym oder ein katalytisch aktives Fragment davon codiert, welches optimale Aktivität im ER oder Golgi bei einem pH zwischen 5,1 und 8,0 hat, wobei die DNA-Bibliothek mindestens zwei DNA-Fragmente, die ein zelluläres Zielsignal-Peptid codieren, und mindestens zwei DNA-Fragmente, die exogene Glycosylierungsenzyme codieren, enthält.
- Vielzahl von Wirtszellen, umfassend eine DNA-Bibliothek nach Anspruch 33.
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US21435800P | 2000-06-28 | 2000-06-28 | |
| US214358P | 2000-06-28 | ||
| US21563800P | 2000-06-30 | 2000-06-30 | |
| US215638P | 2000-06-30 | ||
| US27999701P | 2001-03-30 | 2001-03-30 | |
| US279997P | 2001-03-30 | ||
| PCT/US2001/020553 WO2002000879A2 (en) | 2000-06-28 | 2001-06-27 | Methods for producing modified glycoproteins |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| DE60114830D1 DE60114830D1 (de) | 2005-12-15 |
| DE60114830T2 true DE60114830T2 (de) | 2006-08-03 |
Family
ID=27395978
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| DE60114830T Revoked DE60114830T2 (de) | 2000-06-28 | 2001-06-27 | Verfahren zur herstellung modifizierter glycoproteine |
| DE60139720T Expired - Lifetime DE60139720D1 (de) | 2000-06-28 | 2001-06-27 | Verfahren für die Herstellung modifizierter Glykoproteine |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| DE60139720T Expired - Lifetime DE60139720D1 (de) | 2000-06-28 | 2001-06-27 | Verfahren für die Herstellung modifizierter Glykoproteine |
Country Status (15)
| Country | Link |
|---|---|
| US (13) | US7029872B2 (de) |
| EP (5) | EP2119793A1 (de) |
| JP (2) | JP2004501642A (de) |
| KR (1) | KR100787073B1 (de) |
| AT (2) | ATE309385T1 (de) |
| AU (2) | AU7684201A (de) |
| CA (1) | CA2412701A1 (de) |
| CY (1) | CY1109639T1 (de) |
| DE (2) | DE60114830T2 (de) |
| DK (2) | DK1297172T3 (de) |
| ES (2) | ES2252261T3 (de) |
| MX (1) | MXPA03000105A (de) |
| NZ (1) | NZ523476A (de) |
| PT (1) | PT1522590E (de) |
| WO (1) | WO2002000879A2 (de) |
Families Citing this family (470)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5545553A (en) * | 1994-09-26 | 1996-08-13 | The Rockefeller University | Glycosyltransferases for biosynthesis of oligosaccharides, and genes encoding them |
| PT1137789E (pt) | 1998-12-09 | 2010-10-21 | Phyton Holdings Llc | Um método para produzir glicoproteínas possuindo glicosilação de tipo humano |
| BRPI0015031B1 (pt) | 1999-10-26 | 2017-05-30 | Plant Res Int B V | método para prover uma planta transgênica capaz de expressar um anticorpo com galactosilação terminal e método para obtenção de uma glicoproteína desejada ou fragmento funcional da mesma |
| DK1283265T3 (da) * | 2000-05-17 | 2009-11-09 | Mitsubishi Tanabe Pharma Corp | Fremgangsmåde til fremstilling af protein med reduktion af mannosephosphat i sukkerkæden og glycoprotein fremstillet derved |
| US7625756B2 (en) | 2000-06-28 | 2009-12-01 | GycoFi, Inc. | Expression of class 2 mannosidase and class III mannosidase in lower eukaryotic cells |
| US20050260729A1 (en) * | 2004-03-17 | 2005-11-24 | Hamilton Stephen R | Method of engineering a cytidine monophosphate-sialic acid synthetic pathway in fungi and yeast |
| US7795002B2 (en) | 2000-06-28 | 2010-09-14 | Glycofi, Inc. | Production of galactosylated glycoproteins in lower eukaryotes |
| US8697394B2 (en) | 2000-06-28 | 2014-04-15 | Glycofi, Inc. | Production of modified glycoproteins having multiple antennary structures |
| US20060034828A1 (en) * | 2000-06-28 | 2006-02-16 | Gerngross Tillman U | Immunoglobulins comprising predominantly a GlcNAcMAN5GLCNAC2 glycoform |
| US7598055B2 (en) * | 2000-06-28 | 2009-10-06 | Glycofi, Inc. | N-acetylglucosaminyltransferase III expression in lower eukaryotes |
| US20060034830A1 (en) * | 2000-06-28 | 2006-02-16 | Gerngross Tillman U | Immunoglobulins comprising predominantly a GalGlcNAcMan5GLcNAc2 glycoform |
| KR100787073B1 (ko) * | 2000-06-28 | 2007-12-21 | 글리코파이, 인크. | 변형된 당단백질의 제조방법 |
| US7863020B2 (en) | 2000-06-28 | 2011-01-04 | Glycofi, Inc. | Production of sialylated N-glycans in lower eukaryotes |
| US20060024304A1 (en) * | 2000-06-28 | 2006-02-02 | Gerngross Tillman U | Immunoglobulins comprising predominantly a Man5GlcNAc2 glycoform |
| US20060029604A1 (en) * | 2000-06-28 | 2006-02-09 | Gerngross Tillman U | Immunoglobulins comprising predominantly a GlcNAc2Man3GlcNAc2 glycoform |
| US7449308B2 (en) | 2000-06-28 | 2008-11-11 | Glycofi, Inc. | Combinatorial DNA library for producing modified N-glycans in lower eukaryotes |
| US6803225B2 (en) | 2000-06-30 | 2004-10-12 | Flanders Interuniversity Institute For Biotechnology | Protein glycosylation modification in Pichia pastoris |
| ES2390425T3 (es) | 2000-12-22 | 2012-11-12 | MAX-PLANCK-Gesellschaft zur Förderung der Wissenschaften e.V. | Uso de moléculas de orientación repulsivas (RGM) y sus moduladores |
| US20040214273A1 (en) | 2001-01-19 | 2004-10-28 | Kazuhito Fujiyama | Method for secretory production of glycoprotein having human-type sugar chain using plant cell |
| TWI377253B (en) | 2001-04-16 | 2012-11-21 | Martek Biosciences Corp | Product and process for transformation of thraustochytriales microorganisms |
| US7226903B2 (en) | 2001-10-10 | 2007-06-05 | Neose Technologies, Inc. | Interferon beta: remodeling and glycoconjugation of interferon beta |
| US7173003B2 (en) | 2001-10-10 | 2007-02-06 | Neose Technologies, Inc. | Granulocyte colony stimulating factor: remodeling and glycoconjugation of G-CSF |
| US7157277B2 (en) | 2001-11-28 | 2007-01-02 | Neose Technologies, Inc. | Factor VIII remodeling and glycoconjugation of Factor VIII |
| US7439043B2 (en) * | 2001-10-10 | 2008-10-21 | Neose Technologies, Inc. | Galactosyl nucleotide sugars |
| US8008252B2 (en) * | 2001-10-10 | 2011-08-30 | Novo Nordisk A/S | Factor VII: remodeling and glycoconjugation of Factor VII |
| US7265085B2 (en) * | 2001-10-10 | 2007-09-04 | Neose Technologies, Inc. | Glycoconjugation methods and proteins/peptides produced by the methods |
| US7265084B2 (en) * | 2001-10-10 | 2007-09-04 | Neose Technologies, Inc. | Glycopegylation methods and proteins/peptides produced by the methods |
| US7795210B2 (en) | 2001-10-10 | 2010-09-14 | Novo Nordisk A/S | Protein remodeling methods and proteins/peptides produced by the methods |
| US7696163B2 (en) | 2001-10-10 | 2010-04-13 | Novo Nordisk A/S | Erythropoietin: remodeling and glycoconjugation of erythropoietin |
| US7297511B2 (en) * | 2001-10-10 | 2007-11-20 | Neose Technologies, Inc. | Interferon alpha: remodeling and glycoconjugation of interferon alpha |
| US7214660B2 (en) | 2001-10-10 | 2007-05-08 | Neose Technologies, Inc. | Erythropoietin: remodeling and glycoconjugation of erythropoietin |
| NZ532027A (en) * | 2001-10-10 | 2008-09-26 | Neose Technologies Inc | Remodeling and glycoconjugation of peptides |
| US7399613B2 (en) * | 2001-10-10 | 2008-07-15 | Neose Technologies, Inc. | Sialic acid nucleotide sugars |
| CN105131104B (zh) | 2001-10-10 | 2018-11-16 | 诺和诺德公司 | 肽的重构和糖缀合 |
| US7179617B2 (en) | 2001-10-10 | 2007-02-20 | Neose Technologies, Inc. | Factor IX: remolding and glycoconjugation of Factor IX |
| US7125843B2 (en) | 2001-10-19 | 2006-10-24 | Neose Technologies, Inc. | Glycoconjugates including more than one peptide |
| US7473680B2 (en) * | 2001-11-28 | 2009-01-06 | Neose Technologies, Inc. | Remodeling and glycoconjugation of peptides |
| US20060024292A1 (en) * | 2001-12-27 | 2006-02-02 | Gerngross Tillman U | Immunoglobulins comprising predominantly a Gal2GlcNAc2Man3GlcNAc2 glycoform |
| US20060034829A1 (en) * | 2001-12-27 | 2006-02-16 | Gerngross Tillman U | Immunoglobulins comprising predominantly a MAN3GLCNAC2 glycoform |
| ES2402527T3 (es) | 2001-12-27 | 2013-05-06 | Glycofi, Inc. | Procedimientos para obtener estructuras de carbohidrato de tipo mamífero mediante ingeniería genética |
| US6964784B2 (en) * | 2002-03-07 | 2005-11-15 | Optigenex, Inc. | Method of preparation and composition of a water soluble extract of the bioactive component of the plant species uncaria for enhancing immune, anti-inflammatory, anti-tumor and dna repair processes of warm blooded animals |
| KR101070518B1 (ko) | 2002-03-19 | 2011-10-05 | 스티칭 디엔스트 랜드보위쿤디그 온데조에크 | 식물에서의 gntⅲ 발현 |
| EP2339004B1 (de) * | 2002-03-19 | 2015-02-25 | Stichting Dienst Landbouwkundig Onderzoek | Optimierung von Glycanprozessierung in Pflanzen |
| ATE420198T1 (de) | 2002-04-26 | 2009-01-15 | Kirin Pharma Kk | Methylotrophe hefe,die eine zuckerkette eines säugers herstellt |
| DE60336555D1 (de) | 2002-06-21 | 2011-05-12 | Novo Nordisk Healthcare Ag | Pegylierte glykoformen von faktor vii |
| US7252933B2 (en) * | 2002-06-26 | 2007-08-07 | Flanders Interuniversity Institute For Biotechnology | Protein glycosylation modification in methylotrophic yeast |
| AU2003246286A1 (en) * | 2002-06-29 | 2004-01-19 | Korea Research Institute Of Bioscience And Biotechnology | Hansenula polymorpha mutant strains with defect in outer chain biosynthesis and the production of recombinant glycoproteins using the same strains |
| WO2004028545A1 (en) * | 2002-09-25 | 2004-04-08 | Astrazeneca Ab | A COMBINATION OF A LONG-ACTING β2-AGONIST AND A GLUCOCORTICOSTEROID IN THE TREATMENT OF FIBROTIC DISEASES |
| CL2003002461A1 (es) | 2002-11-27 | 2005-01-07 | Dow Chemical Company Agroscien | Inmunoglobulina que comprende al menos un glicano afucosilado, composicion que la contiene, secuencia nucleotidica y vector que la comprende, procedimiento para producir dicha inmunoglobulina en plantas. |
| ES2574993T3 (es) | 2003-01-22 | 2016-06-23 | Roche Glycart Ag | Construcciones de fusión y uso de las mismas para producir anticuerpos con mayor afinidad de unión al receptor de Fc y función efectora |
| RU2407796C2 (ru) * | 2003-01-22 | 2010-12-27 | Гликарт Биотекнолоджи АГ | КОНСТРУКЦИИ СЛИЯНИЯ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ АНТИТЕЛ С ПОВЫШЕННЫМИ АФФИННОСТЬЮ СВЯЗЫВАНИЯ Fc-РЕЦЕПТОРА И ЭФФЕКТОРНОЙ ФУНКЦИЕЙ |
| DE10303974A1 (de) | 2003-01-31 | 2004-08-05 | Abbott Gmbh & Co. Kg | Amyloid-β(1-42)-Oligomere, Verfahren zu deren Herstellung und deren Verwendung |
| AU2012227297B2 (en) * | 2003-02-20 | 2013-11-14 | Glycofi, Inc. | Combinatorial DNA Library for Producing Modified N-Glycans in Lower Eukaryotes |
| US7332299B2 (en) | 2003-02-20 | 2008-02-19 | Glycofi, Inc. | Endomannosidases in the modification of glycoproteins in eukaryotes |
| KR20060003862A (ko) | 2003-03-14 | 2006-01-11 | 네오스 테크놀로지스, 인크. | 수용성분기폴리머 및 그 접합체 |
| US20070026485A1 (en) | 2003-04-09 | 2007-02-01 | Neose Technologies, Inc. | Glycopegylation methods and proteins/peptides produced by the methods |
| WO2006127896A2 (en) * | 2005-05-25 | 2006-11-30 | Neose Technologies, Inc. | Glycopegylated factor ix |
| US7691603B2 (en) * | 2003-04-09 | 2010-04-06 | Novo Nordisk A/S | Intracellular formation of peptide conjugates |
| US8791070B2 (en) | 2003-04-09 | 2014-07-29 | Novo Nordisk A/S | Glycopegylated factor IX |
| US7932364B2 (en) | 2003-05-09 | 2011-04-26 | Novo Nordisk A/S | Compositions and methods for the preparation of human growth hormone glycosylation mutants |
| WO2005012484A2 (en) | 2003-07-25 | 2005-02-10 | Neose Technologies, Inc. | Antibody-toxin conjugates |
| US9357755B2 (en) * | 2003-10-28 | 2016-06-07 | The University Of Wyoming | Production of human glycosylated proteins in silk worm |
| WO2005042753A1 (en) * | 2003-10-28 | 2005-05-12 | Chesapeake Perl, Inc. | Production of human glycosylated proteins in transgenic insects |
| US20050100965A1 (en) | 2003-11-12 | 2005-05-12 | Tariq Ghayur | IL-18 binding proteins |
| US7507573B2 (en) * | 2003-11-14 | 2009-03-24 | Vib, Vzw | Modification of protein glycosylation in methylotrophic yeast |
| US20080305992A1 (en) | 2003-11-24 | 2008-12-11 | Neose Technologies, Inc. | Glycopegylated erythropoietin |
| EP1694347B1 (de) * | 2003-11-24 | 2013-11-20 | BioGeneriX AG | Glycopegyliertes Erythropoetin |
| US8633157B2 (en) | 2003-11-24 | 2014-01-21 | Novo Nordisk A/S | Glycopegylated erythropoietin |
| US20060040856A1 (en) | 2003-12-03 | 2006-02-23 | Neose Technologies, Inc. | Glycopegylated factor IX |
| US20080318850A1 (en) * | 2003-12-03 | 2008-12-25 | Neose Technologies, Inc. | Glycopegylated Factor Ix |
| US7956032B2 (en) * | 2003-12-03 | 2011-06-07 | Novo Nordisk A/S | Glycopegylated granulocyte colony stimulating factor |
| JP4861830B2 (ja) * | 2003-12-24 | 2012-01-25 | グライコフィ, インコーポレイテッド | 糖タンパク質の産生において、グリカンのマンノシルリン酸化を除去する方法 |
| KR101439880B1 (ko) | 2004-01-08 | 2014-09-12 | 라티오팜 게엠베하 | 펩티드의 오-결합형 글리코실화 |
| KR100604994B1 (ko) | 2004-01-30 | 2006-07-26 | 한국생명공학연구원 | 알파1,6-만노실트랜스퍼라제를 코딩하는 한세눌라폴리모르파 기원의 신규 유전자 및 상기 유전자가 결손된한세눌라 폴리모르파 변이주를 이용한 재조합 당단백질생산 방법 |
| US20050265988A1 (en) * | 2004-03-18 | 2005-12-01 | Byung-Kwon Choi | Glycosylated glucocerebrosidase expression in fungal hosts |
| AU2005233387B2 (en) * | 2004-04-15 | 2011-05-26 | Glycofi, Inc. | Production of galactosylated glycoproteins in lower eukaryotes |
| JP5752582B2 (ja) * | 2004-04-29 | 2015-07-22 | グライコフィ, インコーポレイテッド | 糖タンパク質の作製においてアルファ−マンノシダーゼ抵抗性グリカンを減少させるか又は排除する方法 |
| AU2005238308B8 (en) * | 2004-04-29 | 2009-10-08 | Glycofi, Inc. | Methods for reducing or eliminating alpha-mannosidase resistant glycans in the production of glycoproteins |
| ATE455861T1 (de) | 2004-05-04 | 2010-02-15 | Novo Nordisk Healthcare Ag | O-verknüpfte glykoformen von faktor vii und verfahren zu deren herstellung |
| EP1771066A2 (de) | 2004-07-13 | 2007-04-11 | Neose Technologies, Inc. | Remodellierung von verzweigtem peg und klykolysierung von glucagonähnlichem peptide-1 glp-1 |
| EP1771479A1 (de) * | 2004-07-21 | 2007-04-11 | Glycofi, Inc. | Immunglobuline mit prädominanter gicnacman5glcnac2-glykoform |
| WO2006026992A1 (en) * | 2004-09-07 | 2006-03-16 | Novozymes A/S | Altered structure of n-glycans in a fungus |
| US8268967B2 (en) | 2004-09-10 | 2012-09-18 | Novo Nordisk A/S | Glycopegylated interferon α |
| JP2008514215A (ja) * | 2004-09-29 | 2008-05-08 | ノボ ノルディスク ヘルス ケア アクチェンゲゼルシャフト | 改変タンパク質 |
| ES2572779T3 (es) | 2004-10-29 | 2016-06-02 | Ratiopharm Gmbh | Remodelación y glucopegilación del Factor de Crecimiento de Fibroblastos (FGF) |
| WO2006071856A2 (en) * | 2004-12-23 | 2006-07-06 | Glycofi, Inc. | Immunoglobulins comprising predominantly a man5glcnac2 glycoform |
| ES2449195T3 (es) | 2005-01-10 | 2014-03-18 | Ratiopharm Gmbh | Factor estimulante de colonias de granulocitos glicopegilado |
| DE602006011132D1 (en) * | 2005-03-07 | 2010-01-28 | Plant Res Int Bv | Glycoengineering in pilzen |
| US20060246544A1 (en) * | 2005-03-30 | 2006-11-02 | Neose Technologies,Inc. | Manufacturing process for the production of peptides grown in insect cell lines |
| EP2386571B1 (de) | 2005-04-08 | 2016-06-01 | ratiopharm GmbH | Zusammensetzungen und Verfahren zur Herstellung von Glycosylierungsmutanten eines Proteaseresistenten menschlichen Wachstumshormons |
| GB0510390D0 (en) | 2005-05-20 | 2005-06-29 | Novartis Ag | Organic compounds |
| EP1888098A2 (de) | 2005-05-25 | 2008-02-20 | Neose Technologies, Inc. | Glykopegylierte erythropoetin-formulierungen |
| US7700739B2 (en) * | 2005-06-30 | 2010-04-20 | Abbott Laboratories | IL-12/p40 binding proteins |
| EP2500356A3 (de) | 2005-08-19 | 2012-10-24 | Abbott Laboratories | Immunglobuline mit zweifacher variabler Domäne und ihre Verwendung |
| RU2515108C2 (ru) | 2005-08-19 | 2014-05-10 | Эббви Инк | Иммуноглобулин с двойными вариабельными доменами и его применения |
| US20090215992A1 (en) * | 2005-08-19 | 2009-08-27 | Chengbin Wu | Dual variable domain immunoglobulin and uses thereof |
| US20070105755A1 (en) * | 2005-10-26 | 2007-05-10 | Neose Technologies, Inc. | One pot desialylation and glycopegylation of therapeutic peptides |
| KR20080080081A (ko) * | 2005-08-19 | 2008-09-02 | 네오스 테크놀로지스, 인크. | 글리코페닐화 인자 ⅶ 및 인자 ⅶ에이 |
| US7612181B2 (en) * | 2005-08-19 | 2009-11-03 | Abbott Laboratories | Dual variable domain immunoglobulin and uses thereof |
| JP2009507040A (ja) * | 2005-09-02 | 2009-02-19 | グライコフィ, インコーポレイテッド | 主としてglcnacman3glcnac2糖形態を含む免疫グロブリン |
| AU2005336263A1 (en) * | 2005-09-09 | 2007-03-15 | Glycofi, Inc. | Immunoglobulin comprising predominantly a man7GlcNAc2, Man8GlcNAc2 glycoform |
| EP1934605B1 (de) * | 2005-09-22 | 2014-03-26 | Prosci Incorporated | In hefemutanten produzierte glykosylierte polypeptide und verwendungsverfahren dafür |
| US8906864B2 (en) | 2005-09-30 | 2014-12-09 | AbbVie Deutschland GmbH & Co. KG | Binding domains of proteins of the repulsive guidance molecule (RGM) protein family and functional fragments thereof, and their use |
| KR101105718B1 (ko) * | 2005-10-27 | 2012-01-17 | 한국생명공학연구원 | 돌리칠 포스페이트 만노스 의존알파-1,3-만노실트랜스퍼라제를 코딩하는 한세눌라폴리모르파 기원의 신규 유전자와 상기 유전자가 결손된한세눌라 폴리모르파 변이주를 이용한 재조합 당단백질생산 방법 |
| US20090048440A1 (en) | 2005-11-03 | 2009-02-19 | Neose Technologies, Inc. | Nucleotide Sugar Purification Using Membranes |
| CN101365783B (zh) * | 2005-11-15 | 2013-07-17 | 格利科菲公司 | 具有降低的o-糖基化的糖蛋白的产生 |
| KR101439828B1 (ko) | 2005-11-30 | 2014-09-17 | 애브비 인코포레이티드 | 아밀로이드 베타 단백질에 대한 모노클로날 항체 및 이의 용도 |
| AU2006319358B2 (en) | 2005-11-30 | 2012-01-19 | AbbVie Deutschland GmbH & Co. KG | Anti-Abeta globulomer antibodies, antigen-binding moieties thereof, corresponding hybridomas, nucleic acids, vectors, host cells, methods of producing said antibodies, compositions comprising said antibodies, uses of said antibodies and methods of using said antibodies |
| KR20080094064A (ko) * | 2006-01-17 | 2008-10-22 | 바이오렉스 쎄라퓨틱스, 인코포레이티드 | 식물에서 n-글리칸의 인간화 및 최적화 조성물 및 방법 |
| US20090060921A1 (en) * | 2006-01-17 | 2009-03-05 | Biolex Therapeutics, Inc. | Glycan-optimized anti-cd20 antibodies |
| US20090181041A1 (en) * | 2006-01-23 | 2009-07-16 | Jan Holgersson | Production of proteins carrying oligomannose or human-like glycans in yeast and methods of use thereof |
| US20070190057A1 (en) | 2006-01-23 | 2007-08-16 | Jian Wu | Methods for modulating mannose content of recombinant proteins |
| DK2264060T3 (da) * | 2006-01-26 | 2014-07-28 | Recopharma Ab | Sammensætninger og fremgangsmåder til inhibering af viral adhæsion |
| DK2004819T3 (en) * | 2006-04-05 | 2015-10-05 | Danisco Us Inc | FILAMENTOUS FUNGI WITH REDUCED CONTENT OF UDP-GALACTOFURANOSE |
| AU2007240624A1 (en) * | 2006-04-21 | 2007-11-01 | Wyeth | Methods for high-throughput screening of cell lines |
| EP2032600A2 (de) * | 2006-05-19 | 2009-03-11 | Glycofi, Inc. | Erythropoietinzusammensetzungen |
| WO2007136865A2 (en) * | 2006-05-19 | 2007-11-29 | Glycofi, Inc | Recombinant vectors |
| ATE516346T1 (de) * | 2006-05-24 | 2011-07-15 | Univ Provence Aix Marseille 1 | Herstellung und verwendungen von chimäre glykosyltransferasen mit optimierter glykosylierungsaktivität codierenden gensequenzen |
| AR078117A1 (es) | 2006-06-20 | 2011-10-19 | Protech Pharma S A | Una muteina recombinante del interferon alfa humano glicosilado, un gen que codifica para dicha muteina, un metodo de produccion de dicho gen, un metodo para obtener una celula eucariota productora de dicha muteina, un metodo para producir dicha muteina, un procedimiento para purificar dicha muteina |
| US9187532B2 (en) | 2006-07-21 | 2015-11-17 | Novo Nordisk A/S | Glycosylation of peptides via O-linked glycosylation sequences |
| US7879799B2 (en) * | 2006-08-10 | 2011-02-01 | Institute For Systems Biology | Methods for characterizing glycoproteins and generating antibodies for same |
| ES2902063T3 (es) | 2006-09-08 | 2022-03-24 | Abbvie Bahamas Ltd | Proteínas de unión a interleucina-13 |
| EP2054521A4 (de) | 2006-10-03 | 2012-12-19 | Novo Nordisk As | Verfahren zur reinigung von polypeptid-konjugate |
| MX2009003774A (es) | 2006-10-12 | 2009-04-22 | Genentech Inc | Anticuerpos para linfotoxina-alfa. |
| EP1916259A1 (de) | 2006-10-26 | 2008-04-30 | Institut National De La Sante Et De La Recherche Medicale (Inserm) | Anti-Glycoprotein VI SCFV Fragment zur Thrombosebehandlung |
| WO2008073620A2 (en) * | 2006-11-02 | 2008-06-19 | Neose Technologies, Inc. | Manufacturing process for the production of polypeptides expressed in insect cell-lines |
| US8455626B2 (en) | 2006-11-30 | 2013-06-04 | Abbott Laboratories | Aβ conformer selective anti-aβ globulomer monoclonal antibodies |
| US20090068110A1 (en) * | 2006-12-22 | 2009-03-12 | Genentech, Inc. | Antibodies to insulin-like growth factor receptor |
| FR2912154B1 (fr) | 2007-02-02 | 2012-11-02 | Glycode | Levures genetiquement modifiees pour la production de glycoproteines homogenes |
| PL2148691T3 (pl) | 2007-02-05 | 2015-12-31 | Apellis Pharmaceuticals Inc | Analogi kompstatyny do stosowania w leczeniu stanów zapalnych układu oddechowego |
| WO2008104386A2 (en) | 2007-02-27 | 2008-09-04 | Abbott Gmbh & Co. Kg | Method for the treatment of amyloidoses |
| RU2479629C2 (ru) * | 2007-03-07 | 2013-04-20 | Гликофи, Инк. | Продукция гликопротеинов с модифицированным фукозилированием |
| US8637016B2 (en) | 2007-03-08 | 2014-01-28 | Kalobios Pharmaceuticals, Inc. | EphA3 antibodies for the treatment of solid tumors |
| NZ580245A (en) | 2007-03-22 | 2012-01-12 | Biogen Idec Inc | Binding proteins, including antibodies, antibody derivatives and antibody fragments, that specifically bind cd154 and uses thereof |
| CA2682897C (en) | 2007-04-03 | 2016-11-22 | Biogenerix Ag | Methods of treatment using glycopegylated g-csf |
| WO2008120107A2 (en) | 2007-04-03 | 2008-10-09 | Oxyrane (Uk) Limited | Glycosylation of molecules |
| US20080256056A1 (en) * | 2007-04-10 | 2008-10-16 | Yahoo! Inc. | System for building a data structure representing a network of users and advertisers |
| MX2009011109A (es) | 2007-04-17 | 2009-12-01 | Plant Res Int Bv | Glicosilación tipo mamífera en plantas para la expresión de glicosil-transferasas no mamíferas. |
| RU2549701C2 (ru) | 2007-05-07 | 2015-04-27 | Медиммун, Ллк | Анти-icos антитела и их применение в лечении онкологических, связанных с трансплантацией и аутоиммунных заболеваний |
| EP2068927B1 (de) | 2007-05-14 | 2015-10-21 | MedImmune, LLC | Verfahren zur reduzierung eosinophiler spiegel |
| US20090053167A1 (en) * | 2007-05-14 | 2009-02-26 | Neose Technologies, Inc. | C-, S- and N-glycosylation of peptides |
| US20090232801A1 (en) * | 2007-05-30 | 2009-09-17 | Abbot Laboratories | Humanized Antibodies Which Bind To AB (1-42) Globulomer And Uses Thereof |
| PE20090329A1 (es) * | 2007-05-30 | 2009-03-27 | Abbott Lab | Anticuerpos humanizados contra el globulomero ab(20-42) y sus usos |
| CN101778859B (zh) | 2007-06-12 | 2014-03-26 | 诺和诺德公司 | 改良的用于生产核苷酸糖的方法 |
| US8207112B2 (en) | 2007-08-29 | 2012-06-26 | Biogenerix Ag | Liquid formulation of G-CSF conjugate |
| US8637435B2 (en) * | 2007-11-16 | 2014-01-28 | Merck Sharp & Dohme Corp. | Eukaryotic cell display systems |
| EP2235198B1 (de) | 2007-12-19 | 2018-08-29 | GlycoFi, Inc. | Hefestämme für die Proteinproduktion |
| CA2711307C (en) * | 2008-01-03 | 2020-10-27 | Cornell Research Foundation, Inc. | Glycosylated protein expression in prokaryotes |
| CA2714166A1 (en) * | 2008-02-20 | 2009-08-27 | Glycofi, Inc. | Vectors and yeast strains for protein production |
| US20130189239A1 (en) | 2008-02-27 | 2013-07-25 | Novo Nordisk A/S | Conjugated Factor VIII Molecules |
| US8962803B2 (en) | 2008-02-29 | 2015-02-24 | AbbVie Deutschland GmbH & Co. KG | Antibodies against the RGM A protein and uses thereof |
| CA2715212A1 (en) * | 2008-03-03 | 2009-09-11 | Glycofi, Inc. | Surface display of recombinant proteins in lower eukaryotes |
| CA2717614A1 (en) * | 2008-03-11 | 2009-09-17 | Genentech, Inc. | Antibodies with enhanced adcc function |
| SG190572A1 (en) | 2008-04-29 | 2013-06-28 | Abbott Lab | Dual variable domain immunoglobulins and uses thereof |
| EP2274331B1 (de) | 2008-05-02 | 2013-11-06 | Novartis AG | Verbesserte bindemoleküle auf fibronectin-basis und ihre verwendung |
| MY159667A (en) | 2008-05-09 | 2017-01-13 | Abbvie Inc | Antibodies to receptor of advanced glycation end products (rage) and uses thereof |
| JP2012503471A (ja) * | 2008-05-30 | 2012-02-09 | グライコフィ, インコーポレイテッド | 末端アルファ−1,3−結合ガラクトースを有するタンパク質の製造のための酵母株 |
| CA2726087A1 (en) | 2008-06-03 | 2009-12-10 | Tariq Ghayur | Dual variable domain immunoglobulins and uses thereof |
| RU2010153578A (ru) | 2008-06-03 | 2012-07-20 | Эбботт Лэборетриз (Us) | Иммуноглобулины с двойными вариабельными доменами и их применение |
| TW201014602A (en) | 2008-07-08 | 2010-04-16 | Abbott Lab | Prostaglandin E2 binding proteins and uses thereof |
| RU2011104348A (ru) | 2008-07-08 | 2012-08-20 | Эбботт Лэборетриз (Us) | Иммуноглобулины с двойным вариабельным доменом против простагландина е2 и их применение |
| US8067339B2 (en) | 2008-07-09 | 2011-11-29 | Merck Sharp & Dohme Corp. | Surface display of whole antibodies in eukaryotes |
| EP2331701B1 (de) | 2008-08-08 | 2013-03-20 | Vib Vzw | Glykoproteine mit veränderten glykosylierungsmustern produzierende zellen und verfahren und verwendung davon |
| CN102177247A (zh) | 2008-08-12 | 2011-09-07 | 格利科菲公司 | 用于蛋白生产: ca2+ atp酶过表达的改良的载体和酵母菌株 |
| CA2738028A1 (en) | 2008-09-25 | 2010-04-01 | Glycosyn, Inc. | Compositions and methods for engineering probiotic yeast |
| JP5652206B2 (ja) | 2008-10-01 | 2015-01-14 | 旭硝子株式会社 | 宿主、形質転換体およびその製造方法、ならびにo−グリコシド型糖鎖含有異種蛋白質の製造方法 |
| US20110293605A1 (en) | 2008-11-12 | 2011-12-01 | Hasige Sathish | Antibody formulation |
| BRPI0922807A2 (pt) * | 2008-12-04 | 2015-12-22 | Abbott Lab | imonuglobulinas de domínio variável duplo e usos dos mesmos |
| SG171814A1 (en) | 2008-12-19 | 2011-07-28 | Jennewein Biotechnologie Gmbh | Synthesis of fucosylated compounds |
| CN102239173A (zh) | 2008-12-19 | 2011-11-09 | 动量制药公司 | 与修饰的聚糖相关的方法 |
| WO2010075249A2 (en) | 2008-12-22 | 2010-07-01 | Genentech, Inc. | A method for treating rheumatoid arthritis with b-cell antagonists |
| KR20110110349A (ko) * | 2009-01-29 | 2011-10-06 | 아보트 러보러터리즈 | Il-1 결합 단백질 |
| US20110165063A1 (en) * | 2009-01-29 | 2011-07-07 | Abbott Laboratories | Il-1 binding proteins |
| US8030026B2 (en) * | 2009-02-24 | 2011-10-04 | Abbott Laboratories | Antibodies to troponin I and methods of use thereof |
| CA2751730A1 (en) | 2009-02-25 | 2010-09-02 | Merck Sharp & Dohme Corp. | Metabolic engineering of a galactose assimilation pathway in the glycoengineered yeast pichia pastoris |
| EP2810652A3 (de) | 2009-03-05 | 2015-03-11 | AbbVie Inc. | IL-17-bindende Proteine |
| CN102405237A (zh) | 2009-03-06 | 2012-04-04 | 卡罗拜奥斯制药公司 | 利用EphA3抗体治疗白血病和慢性骨髓增生性疾病 |
| US8283162B2 (en) | 2009-03-10 | 2012-10-09 | Abbott Laboratories | Antibodies relating to PIVKAII and uses thereof |
| EA024695B1 (ru) | 2009-03-16 | 2016-10-31 | Сефалон Острэйлиа Пти Лтд. | Гуманизированные антитела с противоопухолевой активностью |
| TWI548648B (zh) | 2009-03-16 | 2016-09-11 | Dsm智慧財產有限公司 | 於網黏菌門微生物中生產蛋白質之技術 |
| JP5766179B2 (ja) | 2009-04-27 | 2015-08-19 | ノバルティス アーゲー | 筋肉増殖を増加させるための組成物および方法 |
| MX2011011338A (es) | 2009-04-27 | 2012-04-19 | Novartis Ag | Composiciones y metodos de uso para anticuerpos terapeuticos especificos para la subunidad beta1 del receptor de il-12. |
| WO2010138502A2 (en) * | 2009-05-26 | 2010-12-02 | Momenta Pharmaceuticals, Inc. | Production of glycoproteins |
| US8815242B2 (en) * | 2009-05-27 | 2014-08-26 | Synageva Biopharma Corp. | Avian derived antibodies |
| EP2456463A4 (de) * | 2009-07-24 | 2013-12-04 | Merck Sharp & Dohme | Rekombinante ektodomänenexpression von herpes-simplex-virus-glycoproteinen in hefe |
| EP3029070A1 (de) | 2009-08-29 | 2016-06-08 | AbbVie Inc. | Therapeutische dll4-bindende proteine |
| KR20120060877A (ko) | 2009-09-01 | 2012-06-12 | 아보트 러보러터리즈 | 이원 가변 도메인 면역글로불린 및 이의 용도 |
| WO2011029823A1 (en) | 2009-09-09 | 2011-03-17 | Novartis Ag | Monoclonal antibody reactive with cd63 when expressed at the surface of degranulated mast cells |
| WO2011039634A2 (en) | 2009-09-29 | 2011-04-07 | Universiteit Gent | Hydrolysis of mannose-1-phospho-6-mannose linkage to phospho-6-mannose |
| CA2777068C (en) | 2009-10-14 | 2020-05-26 | Kalobios Pharmaceuticals, Inc. | Antibodies to epha3 |
| AU2010306677B2 (en) | 2009-10-15 | 2013-05-23 | Abbvie Inc. | Dual variable domain immunoglobulins and uses thereof |
| RU2012120102A (ru) | 2009-10-16 | 2013-12-10 | Мерк Шарп И Доум Корп. | Способ получения белков в pichia pastoris, у которых отсутствует поддающаяся обнаружению перекрестная связующая активность по отношению к антителам против антигенов клетки-хозяина |
| UY32979A (es) * | 2009-10-28 | 2011-02-28 | Abbott Lab | Inmunoglobulinas con dominio variable dual y usos de las mismas |
| AU2010313492B2 (en) | 2009-10-30 | 2014-05-29 | Merck Sharp & Dohme Corp. | Method for producing therapeutic proteins in Pichia pastoris lacking dipeptidyl aminopeptidase activity |
| WO2011053545A1 (en) * | 2009-10-30 | 2011-05-05 | Merck Sharp & Dohme Corp. | Granulocyte-colony stimulating factor produced in glycoengineered pichia pastoris |
| WO2011051327A2 (en) | 2009-10-30 | 2011-05-05 | Novartis Ag | Small antibody-like single chain proteins |
| US20130011875A1 (en) | 2009-10-30 | 2013-01-10 | Merck Sharpe & Dohme Corp | Methods for the production of recombinant proteins with improved secretion efficiencies |
| WO2011053707A1 (en) | 2009-10-31 | 2011-05-05 | Abbott Laboratories | Antibodies to receptor for advanced glycation end products (rage) and uses thereof |
| WO2011051466A1 (en) | 2009-11-02 | 2011-05-05 | Novartis Ag | Anti-idiotypic fibronectin-based binding molecules and uses thereof |
| CN102740885B (zh) | 2009-11-05 | 2015-04-01 | 梯瓦制药澳大利亚私人有限公司 | 涉及突变的kras或braf基因的癌症的治疗 |
| EP2501805B1 (de) | 2009-11-19 | 2019-02-20 | Oxyrane UK Limited | Hefe stämme die säugetier-ähnliche n-glykane produzieren |
| WO2011068993A1 (en) | 2009-12-02 | 2011-06-09 | Acceleron Pharma Inc. | Compositions and methods for increasing serum half-life of fc fusion proteins. |
| SG181563A1 (en) | 2009-12-08 | 2012-07-30 | Abbott Gmbh & Co Kg | Monoclonal antibodies against the rgm a protein for use in the treatment of retinal nerve fiber layer degeneration |
| CA2785139A1 (en) | 2009-12-22 | 2011-06-30 | Novartis Ag | Tetravalent cd47-antibody constant region fusion protein for use in therapy |
| EA030095B1 (ru) * | 2009-12-28 | 2018-06-29 | ДиЭсЭм АйПи АССЕТС Б.В. | Получение белка гемагглютинин-нейраминидазы в микроводорослях |
| CN102906270B (zh) * | 2009-12-28 | 2016-06-22 | Dsmip资产公司 | 在木糖上生长的重组破囊壶菌和其组合物、制备方法及用途 |
| CA2785971C (en) | 2009-12-28 | 2019-02-12 | Dsm Ip Assets B.V. | Production of heterologous polypeptides in microalgae, microalgal extracellular bodies, compositions, and methods of making and uses thereof |
| WO2011092233A1 (en) | 2010-01-29 | 2011-08-04 | Novartis Ag | Yeast mating to produce high-affinity combinations of fibronectin-based binders |
| CA2788992A1 (en) * | 2010-02-24 | 2011-09-01 | Merck Sharp & Dohme Corp. | Method for increasing n-glycosylation site occupancy on therapeutic glycoproteins produced in pichia pastoris |
| EP2542582A4 (de) | 2010-03-02 | 2013-12-04 | Abbvie Inc | Therapeutische dll4-bindende proteine |
| BR112012025645A2 (pt) | 2010-04-07 | 2017-12-12 | Momenta Pharmaceuticals Inc | glicanos de alta manose. |
| CA2796339C (en) | 2010-04-15 | 2020-03-31 | Abbott Laboratories | Amyloid-beta binding proteins |
| AU2011248625B2 (en) | 2010-04-26 | 2017-01-05 | Pangu Biopharma Limited | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of cysteinyl-tRNA synthetase |
| JP6294074B2 (ja) | 2010-04-27 | 2018-03-14 | エータイアー ファーマ, インコーポレイテッド | イソロイシルtRNA合成酵素のタンパク質フラグメントに関連した治療用、診断用および抗体組成物の革新的発見 |
| WO2011139853A2 (en) | 2010-04-28 | 2011-11-10 | Atyr Pharma, Inc. | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of alanyl trna synthetases |
| CA2797374C (en) | 2010-04-29 | 2021-02-16 | Pangu Biopharma Limited | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of asparaginyl trna synthetases |
| CA2797393C (en) | 2010-04-29 | 2020-03-10 | Atyr Pharma, Inc. | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of valyl trna synthetases |
| US9068177B2 (en) | 2010-04-29 | 2015-06-30 | Atyr Pharma, Inc | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of glutaminyl-tRNA synthetases |
| CA2797978C (en) | 2010-05-03 | 2019-12-03 | Atyr Pharma, Inc. | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of methionyl-trna synthetases |
| CN103096925A (zh) | 2010-05-03 | 2013-05-08 | Atyr医药公司 | 与精氨酰-tRNA合成酶的蛋白片段相关的治疗、诊断和抗体组合物的创新发现 |
| CN103096912A (zh) | 2010-05-03 | 2013-05-08 | Atyr医药公司 | 与苯丙氨酰-α-tRNA合成酶的蛋白片段相关的治疗、诊断和抗体组合物的创新发现 |
| CN102985103A (zh) | 2010-05-04 | 2013-03-20 | Atyr医药公司 | 与p38多-tRNA合成酶复合物相关的治疗、诊断和抗体组合物的创新发现 |
| JP6396656B2 (ja) | 2010-05-14 | 2018-09-26 | エータイアー ファーマ, インコーポレイテッド | フェニルアラニルβtRNA合成酵素のタンパク質フラグメントに関連した治療用、診断用および抗体組成物の革新的発見 |
| SI2571532T1 (sl) | 2010-05-14 | 2017-10-30 | Abbvie Inc. | Proteini, ki vežejo IL-1 |
| EP2576616A4 (de) | 2010-05-27 | 2014-05-21 | Merck Sharp & Dohme | Verfahren zur herstellung von antikörpern mit verbesserten eigenschaften |
| US8962560B2 (en) | 2010-06-01 | 2015-02-24 | Atyr Pharma Inc. | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of Lysyl-tRNA synthetases |
| WO2012006500A2 (en) | 2010-07-08 | 2012-01-12 | Abbott Laboratories | Monoclonal antibodies against hepatitis c virus core protein |
| UY33492A (es) | 2010-07-09 | 2012-01-31 | Abbott Lab | Inmunoglobulinas con dominio variable dual y usos de las mismas |
| CA2804416C (en) | 2010-07-12 | 2020-04-28 | Atyr Pharma, Inc. | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of glycyl-trna synthetases |
| US9120862B2 (en) | 2010-07-26 | 2015-09-01 | Abbott Laboratories | Antibodies relating to PIVKA-II and uses thereof |
| WO2012013823A2 (en) | 2010-07-30 | 2012-02-02 | Glycode | A yeast artificial chromosome carrying the mammalian glycosylation pathway |
| KR20130100118A (ko) | 2010-08-03 | 2013-09-09 | 아비에 인코포레이티드 | 이원 가변 도메인 면역글로불린 및 이의 용도 |
| EP2603526A1 (de) | 2010-08-13 | 2013-06-19 | Medimmune Limited | Monomere polypeptide mit abweichenden fc-regionen und verwendungsverfahren |
| EP3533803B1 (de) | 2010-08-14 | 2021-10-27 | AbbVie Inc. | Anti-amyloid-beta antikörper |
| WO2012022734A2 (en) | 2010-08-16 | 2012-02-23 | Medimmune Limited | Anti-icam-1 antibodies and methods of use |
| HUE058226T2 (hu) | 2010-08-19 | 2022-07-28 | Zoetis Belgium S A | NGF elleni antitestek és alkalmazásuk |
| AU2011293294B2 (en) | 2010-08-25 | 2016-03-24 | Pangu Biopharma Limited | Innovative discovery of therapeutic, diagnostic, and antibody compositions related to protein fragments of Tyrosyl-tRNA synthetases |
| CA2809433A1 (en) | 2010-08-26 | 2012-03-01 | Abbvie Inc. | Dual variable domain immunoglobulins and uses thereof |
| BR112013007263A2 (pt) | 2010-09-29 | 2016-06-14 | Oxyrane Uk Ltd | desmanosilação de n-glicanos fosforilados |
| CA2812870C (en) | 2010-09-29 | 2020-06-09 | Oxyrane Uk Limited | Mannosidases capable of uncapping mannose-1-phospho-6-mannose linkages and demannosylating phosphorylated n-glycans and methods of facilitating mammalian cellular uptake of glycoproteins |
| CN103154037A (zh) | 2010-10-05 | 2013-06-12 | 诺瓦提斯公司 | 抗IL12Rβ1抗体及它们在治疗自身免疫病和炎性疾病中的用途 |
| CN103534392A (zh) | 2010-12-01 | 2014-01-22 | 默沙东公司 | 表面锚定的fc-诱饵抗体展示系统 |
| US9458240B2 (en) | 2010-12-10 | 2016-10-04 | Novartis Pharma Ag | Anti-BAFFR antibody formulations |
| KR20130133247A (ko) | 2010-12-21 | 2013-12-06 | 애브비 인코포레이티드 | Il-1-알파 및 -베타 이특이적 이원 가변 도메인 면역글로불린 및 이의 용도 |
| US20120275996A1 (en) | 2010-12-21 | 2012-11-01 | Abbott Laboratories | IL-1 Binding Proteins |
| DK2673373T6 (da) | 2011-02-08 | 2021-08-23 | Medimmune Llc | Antistoffer, der binder specifikt til staphylococcus aureus alfa-toksin, og fremgangsmåder til anvendelse |
| WO2012115903A2 (en) | 2011-02-25 | 2012-08-30 | Merck Sharp & Dohme Corp. | Yeast strain for the production of proteins with modified o-glycosylation |
| US9170249B2 (en) | 2011-03-12 | 2015-10-27 | Momenta Pharmaceuticals, Inc. | N-acetylhexosamine-containing N-glycans in glycoprotein products |
| JP2014509864A (ja) | 2011-03-23 | 2014-04-24 | グリコド | Gdp−フコースを産生可能な酵母組換え細胞 |
| EP2702077A2 (de) | 2011-04-27 | 2014-03-05 | AbbVie Inc. | Verfahren zur steuerung des galactosylierungsprofil von rekombinant exprimierten proteinen |
| DK2707391T3 (en) | 2011-05-13 | 2018-02-05 | Gamamabs Pharma | ANTIBODIES AGAINST HER3 |
| US9328170B2 (en) | 2011-05-25 | 2016-05-03 | Merck Sharp & Dohme Corp. | Method for preparing Fc containing polypeptides having improved properties |
| JP2014519338A (ja) | 2011-06-16 | 2014-08-14 | ノバルティス アーゲー | 治療薬として使用される可溶性タンパク質 |
| FR2976811A1 (fr) | 2011-06-22 | 2012-12-28 | Lfb Biotechnologies | Utilisation d'un anticorps anti-cd20 a haute adcc pour le traitement de la maladie de waldenstrom |
| CN103747803B (zh) | 2011-06-22 | 2016-10-12 | 国家医疗保健研究所 | 抗axl抗体及其用途 |
| EP2723376B1 (de) | 2011-06-22 | 2018-12-05 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Anti-axl-antikörper und verwendungen damit |
| ES2692519T3 (es) | 2011-07-01 | 2018-12-04 | Novartis Ag | Método para tratar trastornos metabólicos |
| EP2734236A4 (de) | 2011-07-13 | 2015-04-15 | Abbvie Inc | Verfahren und zusammensetzungen zur behandlung von asthma mit anti-il-13-antikörpern |
| KR20140062469A (ko) | 2011-07-20 | 2014-05-23 | 젭테온 인코포레이티드 | 폴리펩티드 분리 방법 |
| WO2013051608A1 (ja) * | 2011-10-03 | 2013-04-11 | 独立行政法人産業技術総合研究所 | 複合型糖鎖加水分解酵素 |
| MX2014004977A (es) | 2011-10-24 | 2014-09-11 | Abbvie Inc | Inmunoaglutinantes dirigidos contra esclerostina. |
| JP2014534218A (ja) | 2011-10-24 | 2014-12-18 | アッヴィ・インコーポレイテッド | Tnfを標的とする免疫結合剤 |
| IN2014CN02963A (de) | 2011-10-28 | 2015-07-03 | Merck Sharp & Dohme | |
| EP3559049A4 (de) | 2011-10-28 | 2019-12-04 | Teva Pharmaceuticals Australia Pty Ltd | Polypeptidkonstrukte und anwendungen davon |
| WO2013066685A1 (en) * | 2011-10-31 | 2013-05-10 | Merck Sharp & Dohme Corp. | Engineered pichia strains with improved fermentation yield and n-glycosylation quality |
| EP2776061B1 (de) | 2011-11-07 | 2019-08-14 | MedImmune, LLC | Multispezifische und multivalente bindungsproteine und verwendungen davon |
| KR101457514B1 (ko) * | 2011-11-21 | 2014-11-03 | 한국생명공학연구원 | 우렁쉥이(멍게) 유래의 시알산 전이효소 및 이를 이용한 시알화 복합당질 합성 방법 |
| CN104114183A (zh) | 2011-12-20 | 2014-10-22 | 印第安纳大学研究及科技有限公司 | 用于治疗糖尿病的基于ctp的胰岛素类似物 |
| TW201333035A (zh) | 2011-12-30 | 2013-08-16 | Abbvie Inc | 針對il-13及/或il-17之雙特異性結合蛋白 |
| EP2617732A1 (de) | 2012-01-19 | 2013-07-24 | Vib Vzw | Werkzeuge und Verfahren zur Expression von Membranproteinen |
| MY176695A (en) | 2012-01-27 | 2020-08-19 | Abbvie Inc | Composition and method for the diagnosis and treatment of diseases associated with neurite degeneration |
| RU2659423C2 (ru) | 2012-02-16 | 2018-07-02 | ЭйТИР ФАРМА, ИНК. | ГИСТИДИЛ-тРНК-СИНТЕТАЗЫ ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ И ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ |
| EP3628326B1 (de) | 2012-03-15 | 2024-02-28 | Oxyrane UK Limited | Verfahren und materialien zur behandlung von morbus pompe |
| US9067990B2 (en) | 2013-03-14 | 2015-06-30 | Abbvie, Inc. | Protein purification using displacement chromatography |
| US9150645B2 (en) | 2012-04-20 | 2015-10-06 | Abbvie, Inc. | Cell culture methods to reduce acidic species |
| US9181572B2 (en) | 2012-04-20 | 2015-11-10 | Abbvie, Inc. | Methods to modulate lysine variant distribution |
| EP2847368B1 (de) | 2012-05-11 | 2020-08-19 | Merck Sharp & Dohme Corp. | Oberflächenverankertes leichtketten-köder-antikörperdisplaysystem |
| US9695454B2 (en) | 2012-05-23 | 2017-07-04 | Glykos Finland Oy | Production of fucosylated glycoproteins |
| EP2856159A4 (de) | 2012-06-01 | 2016-04-13 | Momenta Pharmaceuticals Inc | Verfahren im zusammenhang mit denosumab |
| EP2859018B1 (de) | 2012-06-06 | 2021-09-22 | Zoetis Services LLC | Anti-ngf-antikörper für hunde und verfahren dafür |
| US9216219B2 (en) | 2012-06-12 | 2015-12-22 | Novartis Ag | Anti-BAFFR antibody formulation |
| WO2014004549A2 (en) | 2012-06-27 | 2014-01-03 | Amgen Inc. | Anti-mesothelin binding proteins |
| WO2014011955A2 (en) | 2012-07-12 | 2014-01-16 | Abbvie, Inc. | Il-1 binding proteins |
| EA201590237A1 (ru) | 2012-07-18 | 2015-05-29 | Гликотоп Гмбх | Новые способы лечения антителами против her2 с низкой степенью фукозилирования |
| US9512214B2 (en) | 2012-09-02 | 2016-12-06 | Abbvie, Inc. | Methods to control protein heterogeneity |
| JOP20200308A1 (ar) | 2012-09-07 | 2017-06-16 | Novartis Ag | جزيئات إرتباط il-18 |
| AU2013335065B2 (en) * | 2012-10-22 | 2018-10-25 | Merck Sharp & Dohme Llc | CRZ1 mutant fungal cells |
| JP6407872B2 (ja) | 2012-10-23 | 2018-10-17 | リサーチ コーポレーション テクノロジーズ インコーポレイテッド | 主に均一なグリカン構造を生成するためのピキア・パストリス(Pichiapastoris)株 |
| US20150246118A1 (en) | 2012-10-26 | 2015-09-03 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Lyve-1 antagonists for preventing or treating a pathological condition associated with lymphangiogenesis |
| CA2890263C (en) | 2012-11-01 | 2020-03-10 | Abbvie Inc. | Anti-vegf/dll4 dual variable domain immunoglobulins and uses thereof |
| EP2733153A1 (de) | 2012-11-15 | 2014-05-21 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Verfahren zur Herstellung von Immunkonjugaten und Verwendungen davon |
| WO2014088836A1 (en) | 2012-12-03 | 2014-06-12 | Merck Sharp & Dohme Corp. | O-glycosylated carboxy terminal portion (ctp) peptide-based insulin and insulin analogues |
| US9764006B2 (en) | 2012-12-10 | 2017-09-19 | The General Hospital Corporation | Bivalent IL-2 fusion toxins |
| WO2014090948A1 (en) | 2012-12-13 | 2014-06-19 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Serpin spn4a and biologically active derivatives thereof for use in the treatment of cancer |
| EP2931749B1 (de) | 2012-12-17 | 2019-04-24 | Laboratoire Francais du Fractionnement et des Biotechnologies Societe Anonyme | Verwendung monoklonaler antikörper zur behandlung von entzündungen und bakteriellen infektionen |
| WO2014100139A1 (en) | 2012-12-18 | 2014-06-26 | The Rockefeller University | Glycan-modified anti-cd4 antibodies for hiv prevention and therapy |
| US9550986B2 (en) | 2012-12-21 | 2017-01-24 | Abbvie Inc. | High-throughput antibody humanization |
| US9458244B2 (en) | 2012-12-28 | 2016-10-04 | Abbvie Inc. | Single chain multivalent binding protein compositions and methods |
| US9915665B2 (en) | 2012-12-28 | 2018-03-13 | Abbvie Inc. | High-throughput system and method for identifying antibodies having specific antigen binding activities |
| KR20160002681A (ko) | 2013-01-16 | 2016-01-08 | 인썸(인스티튜트 내셔날 드 라 싼테 에 드 라 리셰르셰메디칼르) | 골격 성장 지연 질환의 예방 또는 치료에 사용하기 위한 용해성 섬유아세포 성장 인자 수용체 3(fgr3) 폴리펩티드 |
| EP2948177A1 (de) | 2013-01-22 | 2015-12-02 | AbbVie Inc. | Verfahren zur optimierung der domänenstabilität von bindungsproteinen |
| SI2953969T1 (sl) | 2013-02-08 | 2020-01-31 | Novartis Ag | Protitelesa proti-IL-17A in njihova uporaba v zdravljenju avtoimunskih in vnetnih motenj |
| EP2956169B1 (de) | 2013-02-12 | 2018-04-11 | THE UNITED STATES OF AMERICA, represented by the S | Monoklonale antikörper zur neutralisierung von noroviren |
| EP2956480B1 (de) | 2013-02-13 | 2019-09-04 | Laboratoire Français du Fractionnement et des Biotechnologies | In hohem masse galactosylierte anti-tnf-alpha-antikörper und verwendungen davon |
| WO2014138188A1 (en) | 2013-03-07 | 2014-09-12 | The General Hospital Corporation | Human ctla4 mutants and use thereof |
| EP2830651A4 (de) | 2013-03-12 | 2015-09-02 | Abbvie Inc | An humanes tnf-alpha bindende humane antikörper und verfahren zur herstellung davon |
| US10017543B2 (en) | 2013-03-13 | 2018-07-10 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Prefusion RSV F proteins and their use |
| WO2014151878A2 (en) | 2013-03-14 | 2014-09-25 | Abbvie Inc. | Methods for modulating protein glycosylation profiles of recombinant protein therapeutics using monosaccharides and oligosacharides |
| JP2016512241A (ja) | 2013-03-14 | 2016-04-25 | アボット・ラボラトリーズAbbott Laboratories | 改良された抗体検出のためのhcvns3組換え抗原およびこの突然変異体 |
| US9017687B1 (en) | 2013-10-18 | 2015-04-28 | Abbvie, Inc. | Low acidic species compositions and methods for producing and using the same using displacement chromatography |
| WO2014158900A1 (en) | 2013-03-14 | 2014-10-02 | Indiana University Research And Technology Corporation | Insulin-incretin conjugates |
| EP2968526A4 (de) | 2013-03-14 | 2016-11-09 | Abbott Lab | Hcv-antigen-antikörper-kombinationstest sowie verfahren und zusammensetzungen zur verwendung darin |
| JP6739329B2 (ja) | 2013-03-14 | 2020-08-12 | アボット・ラボラトリーズAbbott Laboratories | Hcvコア脂質結合ドメインモノクローナル抗体 |
| DK3460054T3 (da) | 2013-03-15 | 2021-01-18 | Atyr Pharma Inc | Histidyl-tRNA-syntetase-Fc-konjugater |
| WO2014149067A1 (en) | 2013-03-15 | 2014-09-25 | Momenta Pharmaceuticals, Inc. | Methods related to ctla4-fc fusion proteins |
| WO2014144280A2 (en) | 2013-03-15 | 2014-09-18 | Abbvie Inc. | DUAL SPECIFIC BINDING PROTEINS DIRECTED AGAINST IL-1β AND / OR IL-17 |
| WO2014173886A1 (en) | 2013-04-22 | 2014-10-30 | Glycotope Gmbh | Anti-cancer treatments with anti-egfr antibodies having a low fucosylation |
| EP2992013B1 (de) | 2013-04-29 | 2019-12-04 | Teva Pharmaceuticals Australia Pty Ltd | Anti-cd38-antikörper und fusionen davon mit abgeschwächtem interferon-alpha-2b |
| US11117975B2 (en) | 2013-04-29 | 2021-09-14 | Teva Pharmaceuticals Australia Pty Ltd | Anti-CD38 antibodies and fusions to attenuated interferon alpha-2B |
| AR096236A1 (es) | 2013-05-09 | 2015-12-16 | The Us Secretary Dept Of Health And Human Services Nat Inst Of Health Office Of Tech Transfer | Anticuerpos de dominio simple vhh dirigidos a norovirus gi.1 y gii.4 y sus usos |
| WO2014186310A1 (en) | 2013-05-13 | 2014-11-20 | Momenta Pharmaceuticals, Inc. | Methods for the treatment of neurodegeneration |
| CN105960414A (zh) | 2013-08-14 | 2016-09-21 | 诺华股份有限公司 | 治疗散发性包涵体肌炎的方法 |
| JP6553042B2 (ja) | 2013-09-05 | 2019-07-31 | ブイアイビー ブイゼットダブリュVib Vzw | 変更されたグリコシル化パターンを有するFc含有分子を産生する細胞並びにその方法及び使用 |
| WO2015044379A1 (en) | 2013-09-27 | 2015-04-02 | INSERM (Institut National de la Santé et de la Recherche Médicale) | A dyrk1a polypeptide for use in preventing or treating metabolic disorders |
| WO2015050959A1 (en) | 2013-10-01 | 2015-04-09 | Yale University | Anti-kit antibodies and methods of use thereof |
| KR102501920B1 (ko) | 2013-10-02 | 2023-02-20 | 메디뮨 엘엘씨 | 중화 항-인플루엔자 a 항체 및 이의 용도 |
| US9598667B2 (en) | 2013-10-04 | 2017-03-21 | Abbvie Inc. | Use of metal ions for modulation of protein glycosylation profiles of recombinant proteins |
| US20160257754A1 (en) | 2013-10-16 | 2016-09-08 | Momenta Pharmaceuticals Inc. | Sialylated glycoproteins |
| US9085618B2 (en) | 2013-10-18 | 2015-07-21 | Abbvie, Inc. | Low acidic species compositions and methods for producing and using the same |
| US9181337B2 (en) | 2013-10-18 | 2015-11-10 | Abbvie, Inc. | Modulated lysine variant species compositions and methods for producing and using the same |
| US10988745B2 (en) | 2013-10-31 | 2021-04-27 | Resolve Therapeutics, Llc | Therapeutic nuclease-albumin fusions and methods |
| CN105849125B (zh) | 2013-11-07 | 2020-05-15 | 国家医疗保健研究所 | 神经调节蛋白变构抗her3抗体 |
| WO2015073884A2 (en) | 2013-11-15 | 2015-05-21 | Abbvie, Inc. | Glycoengineered binding protein compositions |
| FR3016633B1 (fr) | 2014-01-17 | 2018-04-13 | Laboratoire Francais Du Fractionnement Et Des Biotechnologies | Immunoglobuline anti-toxine du charbon |
| WO2015111248A1 (ja) | 2014-01-21 | 2015-07-30 | 学校法人慶應義塾 | 単位dna組成物の調製方法及びdna連結体の作製方法 |
| TW202214691A (zh) | 2014-03-21 | 2022-04-16 | 美商艾伯維有限公司 | 抗-egfr抗體及抗體藥物結合物 |
| TW201622746A (zh) | 2014-04-24 | 2016-07-01 | 諾華公司 | 改善或加速髖部骨折術後身體復原之方法 |
| UA119352C2 (uk) | 2014-05-01 | 2019-06-10 | Тева Фармасьютикалз Острейліа Пті Лтд | Комбінація леналідоміду або помалідоміду і конструкції анти-cd38 антитіло-атенуйований інтерферон альфа-2b та спосіб лікування суб'єкта, який має cd38-експресуючу пухлину |
| EP3161001A2 (de) | 2014-06-25 | 2017-05-03 | Novartis AG | Il17-a spezifische antikörper die mit hyaluronan bindenden peptid-marker fusioniert sind |
| CN113563462A (zh) | 2014-07-15 | 2021-10-29 | 免疫医疗有限责任公司 | 中和抗乙型流感抗体及其用途 |
| AU2015293949B2 (en) | 2014-07-21 | 2019-07-25 | Teknologian Tutkimuskeskus Vtt Oy | Production of glycoproteins with mammalian-like N-glycans in filamentous fungi |
| IL250583B (en) | 2014-08-19 | 2022-07-01 | Merck Sharp & Dohme | Anti-tigit antibodies |
| JO3663B1 (ar) | 2014-08-19 | 2020-08-27 | Merck Sharp & Dohme | الأجسام المضادة لمضاد lag3 وأجزاء ربط الأنتيجين |
| CN108271356A (zh) | 2014-09-24 | 2018-07-10 | 印第安纳大学研究及科技有限公司 | 肠降血糖素-胰岛素缀合物 |
| CN106715473B (zh) | 2014-10-01 | 2021-11-23 | 免疫医疗有限公司 | 替格瑞洛的抗体及其使用方法 |
| AU2015337858B2 (en) | 2014-10-29 | 2020-09-24 | Teva Pharmaceuticals Australia Pty Ltd. | Interferon alpha2b variants |
| WO2016069889A1 (en) | 2014-10-31 | 2016-05-06 | Resolve Therapeutics, Llc | Therapeutic nuclease-transferrin fusions and methods |
| JP2018501814A (ja) | 2014-11-11 | 2018-01-25 | クララ フーズ カンパニー | 卵白タンパク質産生のための方法および組成物 |
| US10072070B2 (en) | 2014-12-05 | 2018-09-11 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Potent anti-influenza A neuraminidase subtype N1 antibody |
| ES2764299T3 (es) | 2014-12-09 | 2020-06-02 | Inst Nat Sante Rech Med | Anticuerpos monoclonales humanos contra AXL |
| US10093733B2 (en) | 2014-12-11 | 2018-10-09 | Abbvie Inc. | LRP-8 binding dual variable domain immunoglobulin proteins |
| WO2016135041A1 (en) | 2015-02-26 | 2016-09-01 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Fusion proteins and antibodies comprising thereof for promoting apoptosis |
| WO2016160976A2 (en) | 2015-03-30 | 2016-10-06 | Abbvie Inc. | Monovalent tnf binding proteins |
| EP3091033A1 (de) | 2015-05-06 | 2016-11-09 | Gamamabs Pharma | Anti-human-her3-antikörper und verwendungen davon |
| SG11201708804WA (en) | 2015-05-07 | 2017-11-29 | Agenus Inc | Anti-ox40 antibodies and methods of use thereof |
| WO2016188911A1 (en) | 2015-05-22 | 2016-12-01 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Human monoclonal antibodies fragments inhibiting both the cath-d catalytic activity and its binding to the lrp1 receptor |
| HUE048284T2 (hu) | 2015-05-29 | 2020-07-28 | Abbvie Inc | Anti-CD40 antitestek és alkalmazásuk |
| HUE056407T2 (hu) | 2015-06-01 | 2022-02-28 | Medimmune Llc | Neutralizáló influenzaellenes kötõmolekulák és alkalmazásaik |
| TW201710286A (zh) | 2015-06-15 | 2017-03-16 | 艾伯維有限公司 | 抗vegf、pdgf及/或其受體之結合蛋白 |
| FR3038517B1 (fr) | 2015-07-06 | 2020-02-28 | Laboratoire Francais Du Fractionnement Et Des Biotechnologies | Utilisation de fragments fc modifies en immunotherapie |
| WO2017005925A1 (en) | 2015-07-09 | 2017-01-12 | Vib Vzw | Cells producing glycoproteins having altered n- and o-glycosylation patterns and methods and use thereof |
| JP2018535655A (ja) | 2015-09-29 | 2018-12-06 | アムジエン・インコーポレーテツド | Asgr阻害剤 |
| MX2018004474A (es) | 2015-10-12 | 2018-11-09 | Aprogen Kic Inc | Anticuerpo anti cd43 y uso del mismo para tratamiento del cancer. |
| JO3555B1 (ar) | 2015-10-29 | 2020-07-05 | Merck Sharp & Dohme | جسم مضاد يبطل فعالية فيروس الالتهاب الرئوي البشري |
| WO2017106129A1 (en) | 2015-12-16 | 2017-06-22 | Merck Sharp & Dohme Corp. | Anti-lag3 antibodies and antigen-binding fragments |
| US11414477B2 (en) | 2016-01-27 | 2022-08-16 | Medimmune, Llc | Methods for preparing antibodies with a defined glycosylation pattern |
| CU24613B1 (es) | 2016-02-06 | 2022-07-08 | Epimab Biotherapeutics Inc | Proteínas de unión a inmunoglobulinas fabs en tandem (fit-ig) biespecíficas que se unen a cmet y egfr |
| RU2018131123A (ru) | 2016-02-17 | 2020-03-17 | Новартис Аг | Антитела к tgf-бета2 |
| KR102640157B1 (ko) | 2016-03-22 | 2024-02-27 | 인쎄름 (엥스띠뛰 나씨오날 드 라 쌍떼 에 드 라 흐쉐르슈 메디깔) | 인간화 항-클라우딘-1 항체 및 이의 용도 |
| CN109563167A (zh) | 2016-06-08 | 2019-04-02 | 艾伯维公司 | 抗b7-h3抗体和抗体药物偶联物 |
| WO2017214335A1 (en) | 2016-06-08 | 2017-12-14 | Abbvie Inc. | Anti-b7-h3 antibodies and antibody drug conjugates |
| MX2018015280A (es) | 2016-06-08 | 2019-08-12 | Abbvie Inc | Conjugados de anticuerpo y farmaco anti-egfr. |
| AU2017277422A1 (en) | 2016-06-08 | 2019-01-03 | Abbvie Inc. | Anti-EGFR antibody drug conjugates |
| WO2017214456A1 (en) | 2016-06-08 | 2017-12-14 | Abbvie Inc. | Anti-cd98 antibodies and antibody drug conjugates |
| CA3027103A1 (en) | 2016-06-08 | 2017-12-14 | Abbvie Inc. | Anti-b7-h3 antibodies and antibody drug conjugates |
| EP4104865A1 (de) | 2016-06-08 | 2022-12-21 | AbbVie Inc. | Anti-egfr-antikörper-wirkstoff-konjugate |
| CN109562170B (zh) | 2016-06-08 | 2023-01-13 | 艾伯维公司 | 抗cd98抗体及抗体药物偶联物 |
| EP3468599A2 (de) | 2016-06-08 | 2019-04-17 | AbbVie Inc. | Anti-cd98-antikörper und antikörperwirkstoffkonjugate |
| MA45554A (fr) | 2016-07-01 | 2019-05-08 | Resolve Therapeutics Llc | Fusions de binucléase optimisées. |
| WO2018014067A1 (en) | 2016-07-19 | 2018-01-25 | Teva Pharmaceuticals Australia Pty Ltd | Anti-cd47 combination therapy |
| BR112019001693A2 (pt) | 2016-07-29 | 2019-07-02 | Ct Hospitalier Universitaire Toulouse | anticorpos direcionados a macrófagos associados a tumores e seus usos |
| NL2017267B1 (en) | 2016-07-29 | 2018-02-01 | Aduro Biotech Holdings Europe B V | Anti-pd-1 antibodies |
| NL2017270B1 (en) | 2016-08-02 | 2018-02-09 | Aduro Biotech Holdings Europe B V | New anti-hCTLA-4 antibodies |
| WO2018026969A2 (en) | 2016-08-03 | 2018-02-08 | Achaogen, Inc. | Plazomicin antibodies and methods of use |
| EP3500301B1 (de) | 2016-08-16 | 2023-08-30 | Epimab Biotherapeutics Inc. | Monovalente asymmetrische tandem-fab-bispezifische antikörper |
| ES2836286T3 (es) | 2016-08-23 | 2021-06-24 | Medimmune Ltd | Anticuerpos anti-VEGF-A y usos de los mismos |
| US10836819B2 (en) | 2016-08-23 | 2020-11-17 | Medimmune Limited | Anti-VEGF-A and anti-ANG2 antibodies and uses thereof |
| EP3512883A1 (de) | 2016-09-13 | 2019-07-24 | Humanigen, Inc. | Epha3-antikörper zur behandlung von lungenfibrose |
| JOP20190055A1 (ar) | 2016-09-26 | 2019-03-24 | Merck Sharp & Dohme | أجسام مضادة ضد cd27 |
| WO2018077926A1 (en) | 2016-10-25 | 2018-05-03 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Monoclonal antibodies binding to the cd160 transmembrane isoform |
| WO2018098370A1 (en) | 2016-11-23 | 2018-05-31 | Immunoah Therapeutics, Inc. | 4-1bb binding proteins and uses thereof |
| CA3046082A1 (en) | 2016-12-07 | 2018-06-14 | Agenus Inc. | Antibodies and methods of use thereof |
| US11471538B2 (en) | 2017-02-10 | 2022-10-18 | INSERM (Institut National de la Santéet de la Recherche Medicale) | Methods and pharmaceutical compositions for the treatment of cancers associated with activation of the MAPK pathway |
| WO2018158398A1 (en) | 2017-03-02 | 2018-09-07 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Antibodies having specificity to nectin-4 and uses thereof |
| WO2018170332A1 (en) * | 2017-03-15 | 2018-09-20 | Nutech Ventures | Extracellular vesicles and methods of using |
| AU2018240117A1 (en) | 2017-03-24 | 2019-09-19 | Beth Israel Deaconess Medical Center, Inc. | Methods for preventing and treating heart disease |
| TWI796329B (zh) | 2017-04-07 | 2023-03-21 | 美商默沙東有限責任公司 | 抗-ilt4抗體及抗原結合片段 |
| AU2018252546A1 (en) | 2017-04-13 | 2019-10-10 | Sairopa B.V. | Anti-SIRPα antibodies |
| US11932694B2 (en) | 2017-04-19 | 2024-03-19 | Bluefin Biomedicine, Inc. | Anti-VTCN1 antibodies and antibody drug conjugates |
| JP2020521780A (ja) | 2017-05-29 | 2020-07-27 | ガママブス ファルマ | 癌関連免疫抑制阻害剤 |
| UY37758A (es) | 2017-06-12 | 2019-01-31 | Novartis Ag | Método de fabricación de anticuerpos biespecíficos, anticuerpos biespecíficos y uso terapéutico de dichos anticuerpos |
| GB201710838D0 (en) | 2017-07-05 | 2017-08-16 | Ucl Business Plc | Bispecific antibodies |
| US11898187B2 (en) | 2017-08-15 | 2024-02-13 | Northwestern University | Protein glycosylation sites by rapid expression and characterization of N-glycosyltransferases |
| SG11202001311VA (en) | 2017-08-22 | 2020-03-30 | Sanabio Llc | Soluble interferon receptors and uses thereof |
| WO2019040780A1 (en) | 2017-08-25 | 2019-02-28 | Five Prime Therapeutics Inc. | ANTI-B7-H4 ANTIBODIES AND METHODS OF USE |
| WO2019075090A1 (en) | 2017-10-10 | 2019-04-18 | Tilos Therapeutics, Inc. | ANTI-LAP ANTIBODIES AND USES THEREOF |
| WO2019148412A1 (en) | 2018-02-01 | 2019-08-08 | Merck Sharp & Dohme Corp. | Anti-pd-1/lag3 bispecific antibodies |
| CA3091174A1 (en) | 2018-02-21 | 2019-08-29 | Five Prime Therapeutics, Inc. | B7-h4 antibody formulations |
| WO2019165075A1 (en) | 2018-02-21 | 2019-08-29 | Five Prime Therapeutics, Inc. | B7-h4 antibody dosing regimens |
| BR112020017925A2 (pt) | 2018-03-02 | 2020-12-22 | Five Prime Therapeutics, Inc. | Anticorpos contra b7-h4 e métodos de uso dos mesmos |
| WO2019177690A1 (en) | 2018-03-12 | 2019-09-19 | Zoetis Services Llc | Anti-ngf antibodies and methods thereof |
| US11530432B2 (en) | 2018-03-19 | 2022-12-20 | Northwestern University | Compositions and methods for rapid in vitro synthesis of bioconjugate vaccines in vitro via production and N-glycosylation of protein carriers in detoxified prokaryotic cell lysates |
| WO2019204346A1 (en) | 2018-04-16 | 2019-10-24 | Northwestern University | METHODS FOR CO-ACTIVATING IN VITRO NON-STANDARD AMINO ACID (nsAA) INCORPORATION AND GLYCOSYLATION IN CRUDE CELLLYSATES |
| CN112888315A (zh) * | 2018-08-21 | 2021-06-01 | 克莱拉食品公司 | 微生物中的蛋白质糖基化的修饰 |
| JP2022500084A (ja) | 2018-09-13 | 2022-01-04 | ザ ボード オブ リージェンツ オブ ザ ユニバーシティー オブ テキサス システム | 新規のlilrb4抗体およびその使用 |
| WO2020058372A1 (en) | 2018-09-19 | 2020-03-26 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Methods and pharmaceutical composition for the treatment of cancers resistant to immune checkpoint therapy |
| JP2022504839A (ja) | 2018-10-10 | 2022-01-13 | ティロス・セラピューティクス・インコーポレイテッド | 抗lap抗体変異体及びその使用 |
| AU2019361923A1 (en) | 2018-10-15 | 2021-06-03 | Five Prime Therapeutics, Inc. | Combination therapy for cancer |
| EP3877413A1 (de) | 2018-11-06 | 2021-09-15 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Verfahren und pharmazeutische zusammensetzungen zur behandlung von akuter myeloischer leukämie durch vernichtung von leukämischen stammzellen |
| JP2022516408A (ja) | 2018-12-14 | 2022-02-28 | インセルム(インスティチュート ナショナル デ ラ サンテ エ デ ラ リシェルシェ メディカル) | 単離されたMHC由来ヒトペプチドおよびCD8+CD45RClowTregの抑制機能を刺激し、かつ活性化するためのその使用 |
| MX2021008081A (es) | 2019-01-04 | 2021-08-05 | Resolve Therapeutics Llc | Tratamiento de la enfermedad de sjogren con proteinas de fusion de nucleasa. |
| WO2020148207A1 (en) | 2019-01-14 | 2020-07-23 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Human monoclonal antibodies binding to hla-a2 |
| JP2022538733A (ja) | 2019-05-20 | 2022-09-06 | インセルム(インスティチュート ナショナル デ ラ サンテ エ デ ラ リシェルシェ メディカル) | 新規抗cd25抗体 |
| BR112021024938A2 (pt) | 2019-06-12 | 2022-01-25 | Novartis Ag | Anticorpos de receptor 1 de peptídeo natriurético e métodos de uso |
| JP2022539879A (ja) | 2019-07-11 | 2022-09-13 | クララ フーズ カンパニー | タンパク質組成物およびその消費可能な製品 |
| WO2021009263A1 (en) | 2019-07-16 | 2021-01-21 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Antibodies having specificity for cd38 and uses thereof |
| US10927360B1 (en) | 2019-08-07 | 2021-02-23 | Clara Foods Co. | Compositions comprising digestive enzymes |
| WO2021067526A1 (en) | 2019-10-02 | 2021-04-08 | Alexion Pharmaceuticals, Inc. | Complement inhibitors for treating drug-induced complement-mediated response |
| EP4037710A1 (de) | 2019-10-04 | 2022-08-10 | Institut National de la Santé et de la Recherche Médicale (INSERM) | Verfahren und pharmazeutische zusammensetzung zur behandlung von eierstockkrebs, brustkrebs oder bauchspeicheldrüsenkrebs |
| WO2021116119A1 (en) | 2019-12-09 | 2021-06-17 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Antibodies having specificity to her4 and uses thereof |
| EP4093760A1 (de) | 2020-01-21 | 2022-11-30 | Yissum Research Development Company of the Hebrew University of Jerusalem Ltd | Verwendung von pflanzenproteinhomologen in kulturmedien |
| CA3167027A1 (en) | 2020-02-05 | 2021-08-12 | Larimar Therapeutics, Inc. | Tat peptide binding proteins and uses thereof |
| US20230272056A1 (en) | 2020-04-09 | 2023-08-31 | Merck Sharp & Dohme Llc | Affinity matured anti-lap antibodies and uses thereof |
| BR112022022045A2 (pt) | 2020-04-30 | 2023-01-10 | Sairopa B V | Anticorpo ou fragmento de ligação ao antígeno do mesmo que se liga ao cd103 humano, um ou mais ácidos nucleicos, sistema de expressão, célula hospedeira, composição, métodos de produção de um anticorpo ou fragmento de ligação ao antígeno, para detectar a presença de cd103 em uma amostra biológica, para tratar ou prevenir uma condição mediada por sinalização de cd103 em um indivíduo em necessidade, para inibir a sinalização de cd103 em uma célula, para inibir a ligação de cd103 à e-caderina presente em uma célula, para retirar as células que expressam cd103 em um indivíduo, para tratar ou prevenir uma doença e agente de imageamento |
| EP4149558A1 (de) | 2020-05-12 | 2023-03-22 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Neues verfahren zur behandlung von kutanen t-zell-lymphomen und tfh-abgeleiteten lymphomen |
| JP2023528235A (ja) | 2020-05-17 | 2023-07-04 | アストラゼネカ・ユーケイ・リミテッド | SARS-CoV-2抗体、並びにこれを選択及び使用する方法 |
| AR122721A1 (es) | 2020-06-25 | 2022-09-28 | Merck Sharp & Dohme | Anticuerpos de alta afinidad dirigidos a tau fosforilada en la serina 413 |
| EP4171614A1 (de) | 2020-06-29 | 2023-05-03 | Resolve Therapeutics, LLC | Behandlung des sjögren-syndroms mit nukleasefusionsproteinen |
| US20230323299A1 (en) | 2020-08-03 | 2023-10-12 | Inserm (Institut National De La Santé Et De La Recherch Médicale) | Population of treg cells functionally committed to exert a regulatory activity and their use for adoptive therapy |
| US20220041694A1 (en) | 2020-08-10 | 2022-02-10 | Astrazeneca Uk Limited | Sars-cov-2 antibodies for treatment and prevention of covid-19 |
| AU2021380966A1 (en) | 2020-11-20 | 2023-06-22 | Alderaan Biotechnology | Anti-cd25 antibodies |
| JP2024504547A (ja) | 2020-11-20 | 2024-02-01 | インセルム(インスティチュート ナショナル デ ラ サンテ エ デ ラ リシェルシェ メディカル) | 抗cd25抗体 |
| WO2022130182A1 (en) | 2020-12-14 | 2022-06-23 | Novartis Ag | Reversal binding agents for anti-natriuretic peptide receptor 1 (npr1) antibodies and uses thereof |
| CN116710482A (zh) | 2021-01-08 | 2023-09-05 | 北京韩美药品有限公司 | 特异性结合4-1bb的抗体及其抗原结合片段 |
| WO2022148414A1 (zh) | 2021-01-08 | 2022-07-14 | 北京韩美药品有限公司 | 特异性结合pd-l1的抗体及其抗原结合片段 |
| WO2022148412A1 (zh) | 2021-01-08 | 2022-07-14 | 北京韩美药品有限公司 | 特异性结合cd47的抗体及其抗原结合片段 |
| WO2022153212A1 (en) | 2021-01-13 | 2022-07-21 | Axon Neuroscience Se | Antibodies neutralizing sars-cov-2 |
| UY39610A (es) | 2021-01-20 | 2022-08-31 | Abbvie Inc | Conjugados anticuerpo-fármaco anti-egfr |
| WO2022200303A1 (en) | 2021-03-23 | 2022-09-29 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Methods for the diagnosis and treatment of t cell-lymphomas |
| EP4320153A1 (de) | 2021-04-09 | 2024-02-14 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Verfahren zur behandlung von anaplastischem grosszelligem lymphom |
| JP2023535884A (ja) | 2021-06-22 | 2023-08-22 | ノバルティス アーゲー | 化膿性汗腺炎の処置における使用のための二特異性抗体 |
| TW202342095A (zh) | 2021-11-05 | 2023-11-01 | 英商阿斯特捷利康英國股份有限公司 | 用於治療和預防covid—19之組成物 |
| EP4186529A1 (de) | 2021-11-25 | 2023-05-31 | Veraxa Biotech GmbH | Verbesserte antikörper-nutzlast-konjugate (aps), hergestellt durch ortsspezifische konjugation mittels genetischer codeerweiterung |
| WO2023094525A1 (en) | 2021-11-25 | 2023-06-01 | Veraxa Biotech Gmbh | Improved antibody-payload conjugates (apcs) prepared by site-specific conjugation utilizing genetic code expansion |
| WO2023110937A1 (en) | 2021-12-14 | 2023-06-22 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Depletion of nk cells for the treatment of adverse post-ischemic cardiac remodeling |
| WO2023144303A1 (en) | 2022-01-31 | 2023-08-03 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Cd38 as a biomarker and biotarget in t-cell lymphomas |
| WO2023187657A1 (en) | 2022-03-30 | 2023-10-05 | Novartis Ag | Methods of treating disorders using anti-natriuretic peptide receptor 1 (npr1) antibodies |
| WO2023198648A1 (en) | 2022-04-11 | 2023-10-19 | Institut National de la Santé et de la Recherche Médicale | Methods for the diagnosis and treatment of t-cell malignancies |
| WO2023198874A1 (en) | 2022-04-15 | 2023-10-19 | Institut National de la Santé et de la Recherche Médicale | Methods for the diagnosis and treatment of t cell-lymphomas |
| CN115386009B (zh) * | 2022-04-26 | 2023-12-01 | 江苏靶标生物医药研究所有限公司 | 一种膜联蛋白v与血管生成抑制剂融合蛋白的构建方法和应用 |
| WO2023209177A1 (en) | 2022-04-29 | 2023-11-02 | Astrazeneca Uk Limited | Sars-cov-2 antibodies and methods of using the same |
| WO2023222886A1 (en) | 2022-05-20 | 2023-11-23 | Depth Charge Ltd | Antibody-cytokine fusion proteins |
| WO2024003310A1 (en) | 2022-06-30 | 2024-01-04 | Institut National de la Santé et de la Recherche Médicale | Methods for the diagnosis and treatment of acute lymphoblastic leukemia |
| EP4309666A1 (de) | 2022-07-21 | 2024-01-24 | Technische Universität Dresden | M-csf zur verwendung bei der prophylaxe und/oder behandlung viraler infektionen bei immunsuppressionszuständen |
| WO2024017998A1 (en) | 2022-07-21 | 2024-01-25 | Technische Universität Dresden | M-csf for use in the prophylaxis and/or treatment of viral infections in states of immunosuppression |
| WO2024018046A1 (en) | 2022-07-22 | 2024-01-25 | Institut National de la Santé et de la Recherche Médicale | Garp as a biomarker and biotarget in t-cell malignancies |
| WO2024023283A1 (en) | 2022-07-29 | 2024-02-01 | Institut National de la Santé et de la Recherche Médicale | Lrrc33 as a biomarker and biotarget in cutaneous t-cell lymphomas |
| WO2024052503A1 (en) | 2022-09-08 | 2024-03-14 | Institut National de la Santé et de la Recherche Médicale | Antibodies having specificity to ltbp2 and uses thereof |
| WO2024079192A1 (en) | 2022-10-12 | 2024-04-18 | Institut National de la Santé et de la Recherche Médicale | Cd81 as a biomarker and biotarget in t-cell malignancies |
| CN115590017B (zh) * | 2022-11-04 | 2023-09-12 | 中国农业大学 | 一种通过降低线粒体温度提高卵母细胞冷冻效果的方法 |
Family Cites Families (115)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US513854A (en) * | 1894-01-30 | Clevis | ||
| US4166329A (en) * | 1978-10-10 | 1979-09-04 | Herbig Charles A | Adjustable arch support for shoes |
| US4414329A (en) | 1980-01-15 | 1983-11-08 | Phillips Petroleum Company | Biochemical conversions by yeast fermentation at high cell densities |
| NZ199722A (en) | 1981-02-25 | 1985-12-13 | Genentech Inc | Dna transfer vector for expression of exogenous polypeptide in yeast;transformed yeast strain |
| US4617274A (en) | 1981-10-29 | 1986-10-14 | Phillips Petroleum Company | Biochemical conversions by yeast fermentation at high cell densities |
| US4775622A (en) | 1982-03-08 | 1988-10-04 | Genentech, Inc. | Expression, processing and secretion of heterologous protein by yeast |
| KR850004274A (ko) | 1983-12-13 | 1985-07-11 | 원본미기재 | 에리트로포이에틴의 제조방법 |
| US4655231A (en) * | 1984-01-09 | 1987-04-07 | Advanced Tobacco Products, Inc. | Snuff and preparation thereof |
| US4837148A (en) | 1984-10-30 | 1989-06-06 | Phillips Petroleum Company | Autonomous replication sequences for yeast strains of the genus pichia |
| US4808537A (en) | 1984-10-30 | 1989-02-28 | Phillips Petroleum Company | Methanol inducible genes obtained from pichia and methods of use |
| US4885242A (en) | 1984-10-30 | 1989-12-05 | Phillips Petroleum Company | Genes from pichia histidine pathway and uses thereof |
| US4855231A (en) | 1984-10-30 | 1989-08-08 | Phillips Petroleum Company | Regulatory region for heterologous gene expression in yeast |
| US4879231A (en) | 1984-10-30 | 1989-11-07 | Phillips Petroleum Company | Transformation of yeasts of the genus pichia |
| US4818700A (en) | 1985-10-25 | 1989-04-04 | Phillips Petroleum Company | Pichia pastoris argininosuccinate lyase gene and uses thereof |
| US5166329A (en) | 1985-10-25 | 1992-11-24 | Phillips Petroleum Company | DNA encoding the alcohol oxidase 2 gene of yeast of the genus Pichia |
| US5032516A (en) | 1985-10-25 | 1991-07-16 | Phillips Petroleum Company | Pichia pastoris alcohol oxidase II regulatory region |
| US4882279A (en) | 1985-10-25 | 1989-11-21 | Phillips Petroleum Company | Site selective genomic modification of yeast of the genus pichia |
| US4935349A (en) | 1986-01-17 | 1990-06-19 | Zymogenetics, Inc. | Expression of higher eucaryotic genes in aspergillus |
| US5837518A (en) * | 1986-01-31 | 1998-11-17 | Genetics Institute, Inc. | Thrombolytic proteins |
| US4812405A (en) | 1986-02-18 | 1989-03-14 | Phillips Petroleum Company | Double auxotrophic mutants of Pichia pastoris and methods for preparation |
| US4925796A (en) | 1986-03-07 | 1990-05-15 | Massachusetts Institute Of Technology | Method for enhancing glycoprotein stability |
| US5272066A (en) | 1986-03-07 | 1993-12-21 | Massachusetts Institute Of Technology | Synthetic method for enhancing glycoprotein stability |
| US4857467A (en) | 1986-07-23 | 1989-08-15 | Phillips Petroleum Company | Carbon and energy source markers for transformation of strains of the genes Pichia |
| US5002876A (en) | 1986-09-22 | 1991-03-26 | Phillips Petroleum Company | Yeast production of human tumor necrosis factor |
| US4683293A (en) | 1986-10-20 | 1987-07-28 | Phillips Petroleum Company | Purification of pichia produced lipophilic proteins |
| US4929555A (en) | 1987-10-19 | 1990-05-29 | Phillips Petroleum Company | Pichia transformation |
| US5135854A (en) | 1987-10-29 | 1992-08-04 | Zymogenetics, Inc. | Methods of regulating protein glycosylation |
| US4816700A (en) * | 1987-12-16 | 1989-03-28 | Intel Corporation | Two-phase non-overlapping clock generator |
| US5004688A (en) | 1988-04-15 | 1991-04-02 | Phillips Petroleum Company | Purification of hepatitis proteins |
| US5047335A (en) | 1988-12-21 | 1991-09-10 | The Regents Of The University Of Calif. | Process for controlling intracellular glycosylation of proteins |
| US5122465A (en) | 1989-06-12 | 1992-06-16 | Phillips Petroleum Company | Strains of pichia pastoris created by interlocus recombination |
| US5032519A (en) | 1989-10-24 | 1991-07-16 | The Regents Of The Univ. Of California | Method for producing secretable glycosyltransferases and other Golgi processing enzymes |
| US5324663A (en) | 1990-02-14 | 1994-06-28 | The Regents Of The University Of Michigan | Methods and products for the synthesis of oligosaccharide structures on glycoproteins, glycolipids, or as free molecules, and for the isolation of cloned genetic sequences that determine these structures |
| US5595900A (en) | 1990-02-14 | 1997-01-21 | The Regents Of The University Of Michigan | Methods and products for the synthesis of oligosaccharide structures on glycoproteins, glycolipids, or as free molecules, and for the isolation of cloned genetic sequences that determine these structures |
| DE4028800A1 (de) | 1990-09-11 | 1992-03-12 | Behringwerke Ag | Gentechnische sialylierung von glykoproteinen |
| CA2058820C (en) | 1991-04-25 | 2003-07-15 | Kotikanyad Sreekrishna | Expression cassettes and vectors for the secretion of human serum albumin in pichia pastoris cells |
| US5962294A (en) | 1992-03-09 | 1999-10-05 | The Regents Of The University Of California | Compositions and methods for the identification and synthesis of sialyltransferases |
| US5602003A (en) | 1992-06-29 | 1997-02-11 | University Of Georgia Research Foundation | N-acetylglucosaminyltransferase V gene |
| EP0726318A1 (de) | 1993-05-14 | 1996-08-14 | The Upjohn Company | Akzeptorpolypeptid für N-Acetylgalaktosaminyltransferase |
| US5484590A (en) | 1993-09-09 | 1996-01-16 | La Jolla Cancer Research Foundation | Expression of the developmental I antigen by a cloned human cDNA encoding a member of a β-1,6-N-acetylglucosaminyltransferase gene family |
| US6300113B1 (en) * | 1995-11-21 | 2001-10-09 | New England Biolabs Inc. | Isolation and composition of novel glycosidases |
| US5683899A (en) | 1994-02-03 | 1997-11-04 | University Of Hawaii | Methods and compositions for combinatorial-based discovery of new multimeric molecules |
| US6069235A (en) * | 1994-02-23 | 2000-05-30 | Monsanto Company | Method for carbohydrate engineering of glycoproteins |
| US6204431B1 (en) | 1994-03-09 | 2001-03-20 | Abbott Laboratories | Transgenic non-human mammals expressing heterologous glycosyltransferase DNA sequences produce oligosaccharides and glycoproteins in their milk |
| PT699237E (pt) * | 1994-03-17 | 2003-07-31 | Merck Patent Gmbh | Fvs de cadeia anti-egfr e anticorpos anti-egfr |
| DE69531686T2 (de) | 1994-06-13 | 2004-07-15 | Banyu Pharmaceutical Co., Ltd. | Glycosyltransferase kodierendes gen und seine verwendung |
| US5545553A (en) | 1994-09-26 | 1996-08-13 | The Rockefeller University | Glycosyltransferases for biosynthesis of oligosaccharides, and genes encoding them |
| WO1996010023A1 (fr) * | 1994-09-27 | 1996-04-04 | Yamanouchi Pharmaceutical Co., Ltd. | Derive de 1,2,3,4-tetrahydroquinoxalindione |
| DE4439759C1 (de) * | 1994-11-07 | 1996-02-01 | Siemens Ag | Photodiodenarray |
| US5849904A (en) | 1994-12-22 | 1998-12-15 | Boehringer Mannheim Gmbh | Isolated nucleic acid molecules which hybridize to polysialyl transferases |
| US5834251A (en) | 1994-12-30 | 1998-11-10 | Alko Group Ltd. | Methods of modifying carbohydrate moieties |
| JP2810635B2 (ja) | 1995-04-03 | 1998-10-15 | 理化学研究所 | 新規糖鎖合成酵素 |
| JPH08336387A (ja) | 1995-06-12 | 1996-12-24 | Green Cross Corp:The | ピキア属酵母由来の糖鎖伸長タンパク及びそのdna |
| EP0874861A1 (de) * | 1995-09-29 | 1998-11-04 | Glycim Oy | Synthetische multivalente slex enthaltende polylactosamine und anwendungsmethode |
| JP3348336B2 (ja) * | 1995-10-26 | 2002-11-20 | 株式会社豊田中央研究所 | 吸着ヒートポンプ |
| US5910570A (en) | 1995-11-13 | 1999-06-08 | Pharmacia & Upjohn Company | Cloned DNA encoding a UDP-GalNAc: polypeptide N-acetylgalactosaminy-ltransferase |
| DE69738177T2 (de) | 1996-08-02 | 2008-07-03 | The Austin Research Institute, Heidelberg | Verbesserte nukleinsäuren, welche für eine chimäre glycosyltransferase kodieren |
| DE69732628T2 (de) | 1996-12-12 | 2005-12-29 | Kirin Beer K.K. | Beta 1-4 n-acetylglucosaminyltransferase sowie das für diese kodierende gen |
| WO2001036432A2 (en) | 1999-11-19 | 2001-05-25 | Human Genome Sciences, Inc. | 18 human secreted proteins |
| US5945314A (en) | 1997-03-31 | 1999-08-31 | Abbott Laboratories | Process for synthesizing oligosaccharides |
| US20040191256A1 (en) | 1997-06-24 | 2004-09-30 | Genentech, Inc. | Methods and compositions for galactosylated glycoproteins |
| US7029870B1 (en) * | 1997-07-03 | 2006-04-18 | Human Genome Sciences, Inc. | Gabaa receptor epsilon subunits |
| AU9073998A (en) * | 1997-09-05 | 1999-03-29 | Glycim Oy | Synthetic divalent slex containing polylactosamines and methods for use |
| JPH11103158A (ja) | 1997-09-26 | 1999-04-13 | Olympus Optical Co Ltd | プリント配線板へのフリップチップ実装方法および実装構造 |
| US7244601B2 (en) | 1997-12-15 | 2007-07-17 | National Research Council Of Canada | Fusion proteins for use in enzymatic synthesis of oligosaccharides |
| JPH11221079A (ja) | 1998-02-04 | 1999-08-17 | Kyowa Hakko Kogyo Co Ltd | 糖転移酵素および該酵素をコードするdna |
| WO1999040208A1 (en) | 1998-02-05 | 1999-08-12 | The General Hospital Corporation | In vivo construction of dna libraries |
| AU3657899A (en) | 1998-04-20 | 1999-11-08 | James E. Bailey | Glycosylation engineering of antibodies for improving antibody-dependent cellular cytotoxicity |
| US6324663B1 (en) * | 1998-10-22 | 2001-11-27 | Vlsi Technology, Inc. | System and method to test internal PCI agents |
| US6870565B1 (en) | 1998-11-24 | 2005-03-22 | Micron Technology, Inc. | Semiconductor imaging sensor array devices with dual-port digital readout |
| PT1137789E (pt) * | 1998-12-09 | 2010-10-21 | Phyton Holdings Llc | Um método para produzir glicoproteínas possuindo glicosilação de tipo humano |
| EP2275540B1 (de) | 1999-04-09 | 2016-03-23 | Kyowa Hakko Kirin Co., Ltd. | Verfahren zur Steuerung der Aktivität von immunologisch funktionellen Molekülen |
| CA2382184A1 (en) * | 1999-08-19 | 2001-03-01 | Kirin Beer Kabushiki Kaisha | Novel yeast mutants and process for producing glycoprotein containing mammalian type sugar chain |
| WO2001025406A1 (en) | 1999-10-01 | 2001-04-12 | University Of Victoria Innovation & Development Corporation | Mannosidases and methods for using same |
| WO2001060860A2 (en) | 2000-02-17 | 2001-08-23 | Millennium Predictive Medicine, Inc. | Genes differentially expressed in human prostate cancer and their use |
| US6410246B1 (en) * | 2000-06-23 | 2002-06-25 | Genetastix Corporation | Highly diverse library of yeast expression vectors |
| US20060029604A1 (en) | 2000-06-28 | 2006-02-09 | Gerngross Tillman U | Immunoglobulins comprising predominantly a GlcNAc2Man3GlcNAc2 glycoform |
| US7625756B2 (en) | 2000-06-28 | 2009-12-01 | GycoFi, Inc. | Expression of class 2 mannosidase and class III mannosidase in lower eukaryotic cells |
| US20060257399A1 (en) | 2000-06-28 | 2006-11-16 | Glycofi, Inc. | Immunoglobulins comprising predominantly a Man5GIcNAc2 glycoform |
| US7449308B2 (en) | 2000-06-28 | 2008-11-11 | Glycofi, Inc. | Combinatorial DNA library for producing modified N-glycans in lower eukaryotes |
| US7795002B2 (en) | 2000-06-28 | 2010-09-14 | Glycofi, Inc. | Production of galactosylated glycoproteins in lower eukaryotes |
| US20060024304A1 (en) | 2000-06-28 | 2006-02-02 | Gerngross Tillman U | Immunoglobulins comprising predominantly a Man5GlcNAc2 glycoform |
| US7863020B2 (en) | 2000-06-28 | 2011-01-04 | Glycofi, Inc. | Production of sialylated N-glycans in lower eukaryotes |
| US20060034830A1 (en) | 2000-06-28 | 2006-02-16 | Gerngross Tillman U | Immunoglobulins comprising predominantly a GalGlcNAcMan5GLcNAc2 glycoform |
| US8697394B2 (en) | 2000-06-28 | 2014-04-15 | Glycofi, Inc. | Production of modified glycoproteins having multiple antennary structures |
| US20050260729A1 (en) | 2004-03-17 | 2005-11-24 | Hamilton Stephen R | Method of engineering a cytidine monophosphate-sialic acid synthetic pathway in fungi and yeast |
| US20060034828A1 (en) | 2000-06-28 | 2006-02-16 | Gerngross Tillman U | Immunoglobulins comprising predominantly a GlcNAcMAN5GLCNAC2 glycoform |
| KR100787073B1 (ko) | 2000-06-28 | 2007-12-21 | 글리코파이, 인크. | 변형된 당단백질의 제조방법 |
| US7598055B2 (en) | 2000-06-28 | 2009-10-06 | Glycofi, Inc. | N-acetylglucosaminyltransferase III expression in lower eukaryotes |
| US6803225B2 (en) | 2000-06-30 | 2004-10-12 | Flanders Interuniversity Institute For Biotechnology | Protein glycosylation modification in Pichia pastoris |
| US7064191B2 (en) * | 2000-10-06 | 2006-06-20 | Kyowa Hakko Kogyo Co., Ltd. | Process for purifying antibody |
| US6946292B2 (en) | 2000-10-06 | 2005-09-20 | Kyowa Hakko Kogyo Co., Ltd. | Cells producing antibody compositions with increased antibody dependent cytotoxic activity |
| JP4655388B2 (ja) | 2001-03-05 | 2011-03-23 | 富士レビオ株式会社 | タンパク質の生産方法 |
| US20050042738A1 (en) | 2001-05-25 | 2005-02-24 | Anita Swarnakar | Carbohydrate-associated proteins |
| NZ592087A (en) * | 2001-08-03 | 2012-11-30 | Roche Glycart Ag | Antibody glycosylation variants having increased antibody-dependent cellular cytotoxicity |
| GB0120311D0 (en) * | 2001-08-21 | 2001-10-17 | Immunoporation Ltd | Treating cells |
| WO2003025148A2 (en) | 2001-09-19 | 2003-03-27 | Nuvelo, Inc. | Novel nucleic acids and polypeptides |
| ES2326964T3 (es) | 2001-10-25 | 2009-10-22 | Genentech, Inc. | Composiciones de glicoproteina. |
| US20060034829A1 (en) | 2001-12-27 | 2006-02-16 | Gerngross Tillman U | Immunoglobulins comprising predominantly a MAN3GLCNAC2 glycoform |
| US20060024292A1 (en) | 2001-12-27 | 2006-02-02 | Gerngross Tillman U | Immunoglobulins comprising predominantly a Gal2GlcNAc2Man3GlcNAc2 glycoform |
| ES2402527T3 (es) | 2001-12-27 | 2013-05-06 | Glycofi, Inc. | Procedimientos para obtener estructuras de carbohidrato de tipo mamífero mediante ingeniería genética |
| US7332299B2 (en) | 2003-02-20 | 2008-02-19 | Glycofi, Inc. | Endomannosidases in the modification of glycoproteins in eukaryotes |
| US7514253B2 (en) | 2003-05-16 | 2009-04-07 | Glycofi, Inc. | URA5 gene and methods for stable genetic integration in yeast |
| EP2581093B1 (de) * | 2003-06-16 | 2015-03-18 | MedImmune, LLC | Influenza-Hämagglutinin- und Neuraminidase-Varianten |
| JP4861830B2 (ja) | 2003-12-24 | 2012-01-25 | グライコフィ, インコーポレイテッド | 糖タンパク質の産生において、グリカンのマンノシルリン酸化を除去する方法 |
| JP2007525146A (ja) * | 2003-12-30 | 2007-09-06 | ガムリンク エー/エス | 生分解性ポリマーを含み且つ分解性を促進させたチューインガム |
| US20050265988A1 (en) | 2004-03-18 | 2005-12-01 | Byung-Kwon Choi | Glycosylated glucocerebrosidase expression in fungal hosts |
| AU2005238308B8 (en) | 2004-04-29 | 2009-10-08 | Glycofi, Inc. | Methods for reducing or eliminating alpha-mannosidase resistant glycans in the production of glycoproteins |
| US7849651B2 (en) * | 2005-05-31 | 2010-12-14 | Kubota Matsushitadenko Exterior Works, Ltd. | Wall materials bracket and insulating wall structure |
| JP2009507040A (ja) | 2005-09-02 | 2009-02-19 | グライコフィ, インコーポレイテッド | 主としてglcnacman3glcnac2糖形態を含む免疫グロブリン |
| CN101365783B (zh) * | 2005-11-15 | 2013-07-17 | 格利科菲公司 | 具有降低的o-糖基化的糖蛋白的产生 |
| US20080274162A1 (en) * | 2007-05-04 | 2008-11-06 | Jeffrey Nessa | Method, composition, and delivery mode for treatment of prostatitis and other urogenital infections using a probiotic rectal suppository |
| TW201028431A (en) * | 2008-10-31 | 2010-08-01 | Lonza Ag | Novel tools for the production of glycosylated proteins in host cells |
| JP2011039027A (ja) * | 2009-07-14 | 2011-02-24 | Pacific Ind Co Ltd | 金属調樹脂カバー及びその製造方法並びに車両用ドアハンドル |
| US9439972B2 (en) * | 2012-09-10 | 2016-09-13 | Ad Lunam Labs, Inc. | Antifungal serum |
-
2001
- 2001-06-27 KR KR1020027017911A patent/KR100787073B1/ko not_active IP Right Cessation
- 2001-06-27 DK DK01954606T patent/DK1297172T3/da active
- 2001-06-27 MX MXPA03000105A patent/MXPA03000105A/es active IP Right Grant
- 2001-06-27 DK DK04025648T patent/DK1522590T3/da active
- 2001-06-27 EP EP09004758A patent/EP2119793A1/de not_active Withdrawn
- 2001-06-27 AU AU7684201A patent/AU7684201A/xx active Pending
- 2001-06-27 DE DE60114830T patent/DE60114830T2/de not_active Revoked
- 2001-06-27 CA CA002412701A patent/CA2412701A1/en not_active Abandoned
- 2001-06-27 ES ES01954606T patent/ES2252261T3/es not_active Expired - Lifetime
- 2001-06-27 DE DE60139720T patent/DE60139720D1/de not_active Expired - Lifetime
- 2001-06-27 US US09/892,591 patent/US7029872B2/en not_active Expired - Fee Related
- 2001-06-27 JP JP2002506194A patent/JP2004501642A/ja active Pending
- 2001-06-27 EP EP10011297A patent/EP2322644A1/de not_active Withdrawn
- 2001-06-27 EP EP01954606A patent/EP1297172B1/de not_active Revoked
- 2001-06-27 AU AU2001276842A patent/AU2001276842B2/en not_active Ceased
- 2001-06-27 PT PT04025648T patent/PT1522590E/pt unknown
- 2001-06-27 EP EP10012383.5A patent/EP2339013B1/de not_active Expired - Lifetime
- 2001-06-27 ES ES04025648T patent/ES2330330T3/es not_active Expired - Lifetime
- 2001-06-27 NZ NZ523476A patent/NZ523476A/en not_active IP Right Cessation
- 2001-06-27 AT AT01954606T patent/ATE309385T1/de not_active IP Right Cessation
- 2001-06-27 AT AT04025648T patent/ATE440959T1/de active
- 2001-06-27 EP EP04025648A patent/EP1522590B1/de not_active Revoked
- 2001-06-27 WO PCT/US2001/020553 patent/WO2002000879A2/en active IP Right Grant
-
2005
- 2005-09-30 US US11/240,432 patent/US7326681B2/en not_active Expired - Fee Related
- 2005-10-11 US US11/249,061 patent/US7923430B2/en not_active Expired - Fee Related
- 2005-11-01 US US11/265,444 patent/US20070178551A1/en not_active Abandoned
- 2005-11-10 US US11/271,217 patent/US20070105127A1/en not_active Abandoned
- 2005-11-10 US US11/271,235 patent/US7629163B2/en not_active Expired - Fee Related
-
2007
- 2007-10-30 US US11/981,408 patent/US8211691B2/en not_active Expired - Fee Related
-
2009
- 2009-08-27 US US12/549,062 patent/US7981660B2/en not_active Expired - Fee Related
- 2009-11-25 CY CY20091101214T patent/CY1109639T1/el unknown
-
2010
- 2010-10-11 US US12/901,843 patent/US20110020870A1/en not_active Abandoned
-
2011
- 2011-03-17 JP JP2011059620A patent/JP2011167194A/ja active Pending
-
2012
- 2012-05-01 US US13/461,111 patent/US20120322100A1/en not_active Abandoned
-
2013
- 2013-07-03 US US13/934,551 patent/US20130295604A1/en not_active Abandoned
-
2014
- 2014-05-07 US US14/271,475 patent/US20140234902A1/en not_active Abandoned
-
2015
- 2015-10-30 US US14/927,519 patent/US20160068880A1/en not_active Abandoned
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| DE60114830T2 (de) | Verfahren zur herstellung modifizierter glycoproteine | |
| AU2001276842A1 (en) | Methods for producing modified glycoproteins | |
| AU2001276842A2 (en) | Methods for producing modified glycoproteins | |
| US8445227B2 (en) | N-acetylglucosaminyltransferase III expression in lower eukaryotes | |
| US20150051381A1 (en) | Combinatorial dna library for producing modified n-glycans in lower eukaryotes | |
| CN101484585A (zh) | 在低等真核生物中生产唾液酸化n-聚糖 | |
| JP2010220616A (ja) | 下等真核生物細胞におけるクラス2マンノシダーゼおよびクラスiiiマンノシダーゼの発現 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 8363 | Opposition against the patent | ||
| 8331 | Complete revocation |