-
HINTERGRUND DER ERFINDUNG
-
Glycosylierungswege
-
Neu
synthetisierte Proteine können
in Zellen einer weiteren Prozessierung unterliegen, die als posttranslationale
Modifikaton bekannt ist. Im Besonderen können Zuckerreste enzymatisch
hinzugefügt
werden, ein Prozess, der als Glycosylierung bekannt ist. Die resultierenden
Proteine, die kovalent gekoppelte Oligosaccharid-Seitenketten tragen,
sind als glycosylierte Proteine oder Glycoproteine bekannt. Bakterien
glycosylieren Proteine im Allgemeinen nicht; in Zellen, in denen
Glycosylierung vorkommt, tritt sie im Allgemeinen an unspezifischen
Stellen in dem Protein auf (Moens and Vanderleyden, Arch. Microbiol.
1997 168 (3): 169–175).
-
Eukaryonten
hängen
im Allgemeinen ein spezifisches Oligosaccharid an die Seitenkette
eines Protein-Asparaginrests an, im Besonderen an ein Asparagin,
das in der Sequenz Asn-Xaa-Ser/Thr/Cys vorkommt (wobei Xaa eine
beliebige Aminosäure
darstellt). Nach der Anheftung der Saccharid-Einheit, die als ein
N-Glycan bekannt
ist, können
in vivo weitere Modifikationen auftreten. Im Allgemeinen treten
diese Modifikationen in einer geordneten Folge enzymatischer Reaktionen
auf, die als eine Kaskade bekannt sind. Verschiedene Organismen
stellen unterschiedliche Glycosylierungs-Enzyme (Glycosyltransferasen
und Glycosidasen) und verschiedene Glycosyl-Substrate bereit, sodass
die endgültige
Zusammensetzung einer Zucker-Seitenkette in Abhängigkeit von dem Wirt ausgesprochen
stark variieren kann.
-
Mikroorganismen
wie filamentöse
Pilze und Hefen (niedere Eukaryonten), hängen zum Beispiel im Allgemeinen
zusätzliche
Mannose- und/oder Mannosylphosphat-Zucker an. Das resultierende
Glycan ist als "High-Mannose"-Typ oder als ein
Mannan bekannt. Im Gegensatz dazu kann die im Entstehen begriffene
Oligosaccharid-Seitenkette in tierischen Zellen zurechtgetrimmt
werden, wobei einige Mannosereste entfernt werden und die Kette
mit zusätzlichen
Zuckerresten verlängert
wird, die im Allgemeinen in den N-Glycanen von niederen Eukaryonten
nicht auftreten. Siehe R. K. Bretthauer, et al. Biotechnology and
Applied Biochemistry,; 1999, 30, 193–200; W. Martinet, et al. Biotechnology
Letters, 1998, 20, 1171–1177;
S. Weikert, et al. Nature Biotechnology Letters, 1999, 17, 1116–1121; M.
Malissard, et al. Biochemical and Biophysical Research Communications,
2000, 267, 169–173;
Jarvis, et al. 1998 Engineering N-glycosylation pathways in the
baculovirus-insect
cell system, Current Opinion in Biotechnology, 9: 528–533; und
M. Takeuchi, 1997 Trends in Glycoscience and Glycotechnology, 1997,
9, S29–S35.
-
Die
N-Glycane, die in Menschen und Tieren hergestellt werden, werden
im Allgemeinen als komplexe N-Glycane bezeichnet. Ein komplexes
N-Glycan bedeutet eine Struktur mit im Allgemeinen zwei bis sechs äußeren Verzweigungen,
wobei eine Sialyl-lactosamin-Sequenz an die innere Kernstruktur
Man3GlcNAc2 gekoppelt
ist. Ein komplexes N-Glycan hat mindestens eine Verzweigung, und
vorzugsweise mindestens zwei, von sich abwechselnden GlcNAc- und
Galactose (Gal)-Resten, die in Oligosacchariden enden, wie zum Beispiel: NeuNAc-;
NeuAcα2-6GalNAcα1-; NeuAcα2-3Ga1β1-3GalNAcα1-; NeuAcα2-3/6Galβ1-4GlcNAcβ1-; GlcNAcα1-4Ga1β1-(ausschließlich Mucine);
Fucα1-2Ga1β1-(Blutgruppe
H). An Galactose-, GalNAc- und GlcNAc-Resten können Sulfatester auftreten
und an Mannoseresten können
Phosphatester auftreten. NeuAc (Neu: Neuraminsäure; Ac: Acetylrest) kann O-acetyliert oder durch
NeuGl (N-Glycolylneuraminsäure)
ausgetauscht sein. Komplexe N-Glycane können auch innerhalb der Kette
Substitutionen aus zweiteilenden GlcNAc und Kern-Fucose (Fuc) tragen.
-
Glycosylierung
beim Menschen beginnt mit einem aufeinanderfolgenden Satz von Reaktionen
im endoplasmatischen Retikulum (ER), was zu einer Kern-Oligosaccharid-Struktur
führt,
die an den Asparaginrest in der Sequenz Asn-Xaa-Ser/Thr von neu synthetisierten Proteinen übertragen
wird (siehe 1A). Im ER tritt eine weitere
Prozessierung durch Glucosidasen und Mannosidasen auf, bevor das
im Entstehen begriffene Glycoprotein in den frühen Golgi-Apparat übertragen
wird, wo zusätzliche
Mannosereste durch Golgi-spezifische 1,2-Mannosidasen entfernt werden.
Die Prozessierung wird fortgesetzt, während das Protein durch den Golgi
voranschreitet. In dem mittleren Golgi fügt eine Vielzahl modifizierender
Enzyme, beinhaltend N-Acetylglucosamin-Transferasen (GnTI, GnTII,
GnTIII, GnTIV, GnTV, GnTVI), Mannosidase II, Fucosyltransferasen, spezifische
Zuckerreste and und entfernt diese (siehe 1B). Schließlich wirken
in dem Trans-Golgi Galactosyltransferasen und Sialyltransferasen
(ST) auf die N-Glycane und das fertiggestellte Glycoprotein wird
aus dem Golgi-Apparat freigesetzt. Die Protein-N-Glycane von tierischen Glycoproteinen
haben bi-, tri-, oder tetra-antennäre Strukturen und können im
Allgemeinen Galactose, Fucose und N-Acetylglucosamin beinhalten. Im
Allgemeinen bestehen die terminalen Reste der N-Glycane aus Sialinsäure. Eine
typische Struktur eines menschlichen N-Glycans ist in 1B gezeigt.
-
Zucker-Nucleotid-Vorläufer
-
Die
N-Glycane tierischer Glycoproteine beinhalten im Allgemeinen Galactose,
Fucose und terminale Sialinsäure.
Diese Zucker werden im Allgemeinen nicht an Glycoproteinen gefunden,
die in Hefe und filamentösen
Pilzen hergestellt werden. In Menschen wird im Allgemeinen der gesamte
Bereich an Nucleotid-Zucker-Vorläufer
(z.B. UDP-N-Acetylglucosamin, UDP-N-Acetylgalactosamin, CMP-N-Acetylneuraminsäure, UDP-Galactose,
GDP-Fucose, etc.) im Cytosol synthetisiert und in den Golgi transportiert,
wo sie durch Glycosyltransferasen an das Kern-Oligosaccharid angeheftet werden (Sommers
and Hirschberg, 1981, J. Cell Biol. 91 (2): A409–A406; Sommers and Hirschberg
1982 J. Biol. Chem. 257 (18): 811–817; Perez and Hirschberg 1987
Methods in Enzymology 138: 709–715).
-
Glycosyl-Transfer-Reaktionen
bringen in der Regel ein Nebenprodukt hervor, bei dem es sich um
ein Nucleosid-Diphosphat oder -Monophosphat handelt. Während Monophosphate
im Austausch gegen Nucleosid-Triphosphat-Zucker durch einen Antiport-Mechanismus
direkt exportiert werden können,
müssen
Diphospho-Nucleoside
(z.B. GDP) durch Phosphatasen (z.B. GDPase) gespalten werden, wobei
Nucleosid-Monophosphate und anorganisches Phosphat entstehen, bevor
diese exportiert werden. Diese Reaktion ist für eine wirkungsvolle Glycosylierung
wichtig; es zeigte sich zum Beispiel, dass GDPase aus S. cerevisiae
für die
Mannosylierung notwendig ist. Die GDPase hat jedoch gegenüber UDP
eine um 90% reduzierte Aktivität
(Berninsone et al., 1994 J. Biol. Chem. 269 (1): 207–211α). Niederen
Eukaryonten fehlt im Allgemeinen UDP-spezifische Diphosphatase-Aktivität im Golgi,
da diese für
die Golgi-basierte Glycoprotein-Synthese keine UDP-Zucker-Vorläufer verwenden.
Man fand heraus, dass Schizosaccharomyces pombe, eine Hefe, von
der sich zeigte, dass sie Galactose-Reste an Zellwand-Polysaccharide
(aus UDP-Galactose) anfügt,
spezifische UDPase-Aktivität
besitzt, was die Notwendigkeit eines solchen Enzyms anzeigt (Berninsone
et al., 1994). UDP ist als ein starker Hemmstoff von Glycosyltransferasen
bekannt und das Entfernen dieses Glycosylierungs-Nebenprodukts ist
wichtig, um in dem Lumen des Golgi Glycosyltransferase-Hemmung zu
verhindern (Khatara et al., 1974). Siehe Berninsone, P., et al.,
1995 J. Biol. Chem. 270 (24): 14564–14567; Beaudet, L. et al.
1998 Abc Transporters: Biochemical, Cellular, and Molecular Aspects.
191: 397–413.
-
Kompartimentierung von
Glycosylierungs-Enzymen
-
Glycosyltransferasen
und Mannosidasen kleiden die innere (luminale) Oberfläche des
ER und des Golgi-Apparates aus und stellen dabei eine katalytische
Oberfläche
bereit, die die sequenzielle Prozessierung von Glycoproteinen erlaubt,
während
diese durch das ER und das Golgi-Netzwerk fortschreiten. Die zahlreichen
Kompartimente des Cis-, des mittleren und des Trans-Golgi und das
Trans-Golgi-Netzwerk
(TGN) stellen die verschiedenen Lokalitäten bereit, in denen die geordnete
Abfolge von Glycosylierungs-Reaktionen stattfinden kann. Sobald
ein Glycoprotein von der Synthese im ER bis hin zu seiner vollen
Reife im späten
Golgi oder TGN fortschreitet, wird es sequenziell verschiedenen
Glycosidasen, Mannosidasen und Glycosyltransferasen ausgesetzt,
sodass eine spezifische N-Glycan-Struktur synthetisiert werden kann.
Die Enzyme beinhalten im Allgemeinen eine katalytische Domäne, eine
Stammregion, eine Membran-durchdringende Region und einen N-terminalen cytoplasmatischen
Schwanz. Die letzteren drei strukturellen Komponenten sind für das Lenken eines
Glycosylierungs-Enzyms zu einem geeigneten Ort verantwortlich.
-
Lokalisierungs-Sequenzen
von einem Organismus können
auch in anderen Organismen funktionieren. Von der Membran-durchdringenden
Region einer α-2,6-Sialyltransferase
(α-2,6-ST)
aus Ratten zum Beispiel, ein Enzym, von dem man weiß, dass
es in dem Trans-Golgi der Ratte lokalisiert ist, wurde gezeigt,
dass es auch für
die Lokalisierung eines Reportergens (Invertase) in dem Hefe-Golgi
verantwortlich ist (Schwientek, et al., 1995). Genau die gleiche
Membran-durchdringende
Region wurde jedoch als Teil einer α-2,6-Sialyltransferase von vollständiger Länge in dem
ER zurückgehalten
und nicht weiter in den Golgi von Hefe transportiert (Krezdorn et
al., 1994). Ein menschliches GalT von vollständiger Länge wurde trotz nachgewiesener
hoher Transkriptionsspiegel in Hefe nicht einmal synthetisiert.
Andererseits war die Transmembran-Region des gleichen menschlichen
GalT, nachdem sie an einen Invertase-Reporter fusioniert worden
war, in der Lage die Lokalisation hin zum Hefe-Golgi auszurichten,
wenn auch in niedrigen Produktionsspiegeln. Schwientek und Mitarbeiter
zeigten, dass die Fusion von 28 Aminosäuren einer Hefe-Mannosyltransferase
(Mnt1), ein Bereich, umfassend einen N-terminalen cytoplasmatischen
Schwanz, eine Transmembran-Region
und acht Aminosäuren
der Stamm-Region, mit der katalytischen Domäne von menschlichem GalT für die Lokalisation
eines aktiven GalT im Golgi ausreichend ist (Schwientek et al. 1995
J. Biol. Chem. 270 (10): 5483–5489).
Andere Galactosyltransferasen scheinen sich auf Wechselwirkungen
mit Enzymen zu stützen,
die in bestimmten Organellen ansässig
sind, da sie nach Entfernen ihrer Transmembran-Region noch zur richtigen
Lokalisation in der Lage sind.
-
Die
falsche Lokalisation eines Glycosylierungs-Enzyms kann das richtige
Funktionieren des Enzyms im Stoffwechselweg verhindern. Aspergillus
nidulans, zum Beispiel, der zahlreiche α-1,2-Mannosidasen besitzt (Eades
and Hintz, 2000 Gene 255 (1): 25–34), fügt kein GlcNAc an Man5GlcNAc2 an, wenn
er mit dem Kaninchen GnTI-Gen transformiert wurde, trotz eines hohen
allgemeinen Spiegels an GnTI-Aktivität (Kalsner et al., 1995). GnTI,
obwohl in aktiver Form exprimiert, kann unkorrekt lokalisiert sein,
derart, dass das Enzym mit beiden seiner Substrate keinen Kontakt
bekommt: das im Entstehen befindliche N-Glycan des Glycoproteins
und UDP-GlcNAc. Alternativ kann es möglich sein, dass der Wirtsorganismus
die angemessenen Spiegel an UDP-GlcNAc in dem Golgi nicht bereitstellt.
-
Glycoproteine, die therapeutisch
verwendet werden
-
Ein
wesentlicher Anteil der Proteine, die aus Menschen oder anderen
Tieren isoliert werden, sind glycosyliert. Unter den Proteinen,
die therapeutisch verwendet werden, sind etwa 70% glycosyliert.
Wenn jedoch ein therapeutisches Protein in einem Mikroorganismus
wie Hefe hergestellt wird und unter Verwendung des endogenen Stoffwechselwegs
glycosyliert wird, ist seine therapeutische Wirksamkeit im Allgemeinen
stark reduziert. Solche Glycoproteine sind im Allgemeinen in Menschen
immunogen und zeigen nach der Verabreichung eine verkürzte in
vivo-Halbwertszeit
(Takeuchi, 1997).
-
Spezifische
Rezeptoren in Menschen und Tieren können terminale Mannose-Reste erkennen und
die rasche Beseitigung des Proteins aus dem Blutstrom verstärken. Zusätzliche
nachteilige Wirkungen können Veränderungen
in der Faltung, der Löslichkeit,
der Zugänglichkeit
für Proteasen,
dem Verkehr, dem Transport, der Kompartimentierung, der Sekretion,
der Erkennung durch andere Proteine oder Faktoren, der Antigenität oder der
Allergenität
des Proteins beinhalten. Entsprechend war es notwendig therapeutische
Glycoproteine in tierischen Wirtsystemen herzustellen, derart, dass
das Muster an Glycosylierung identisch oder zumindest ähnlich dem
in Menschen oder zu den beabsichtigten Empfänger-Arten ist. In den meisten
Fällen
wird ein Säuger-Wirtsystem
wie die Säuger-Zellkultur
verwendet.
-
Systeme für die Herstellung
therapeutischer Glycoproteine
-
Um
therapeutische Proteine herzustellen, die geeignete Glycoformen
haben und zufriedenstellende therapeutische Wirkungen besitzen,
sind Expressionssysteme verwendet worden, die auf Tieren oder Pflanzen
beruhen. Die erhältlichen
Systeme beinhalten:
- 1. Ovar-Zellen des chinesischen
Hamsters (CHO), Maus-Fibroblasten-Zellen und Maus-Myolom-Zellen (Arzneimittelforschung.
1998 Aug; 48 (8): 870–880);
- 2. transgene Tiere wie Ziegen, Schafe, Mäuse und andere (Dente Prog.
Clin. Biol. 1989 Res. 300: 85–98, Ruther
et al., 1988 Cell 53 (6): 847–856;
Ware, J., et al. 1993 Thrombosis and Haemostasis 69 (6): 1194–1194; Cole,
E. S., et al. 1994 J. Cell Biochem. 265–265);
- 3. Pflanzen (Arabidopsis thaliana, Tabak etc.) (Staub, et al.
2000 Nature Biotechnology 18 (3): 333–338) (McGarvey, P. B., et
al. 1995 Bio-Technology
13 (13): 1484–1487;
Bardor, M., et al. 1999 Trends in Plant Science 4 (9): 376–380);
- 4. Insekten-Zellen (Spodoptera frugiperda Sf9, Sf21, Trichoplusia
ni, etc. in Kombination mit rekombinanten Baculoviren, wie multiples
nukleäres
Polyeder-Virus von Autographa californica (das Lepidoptera-Zellen
infiziert) (Altmans et al., 1999 Glycoconj. J. 16 (2): 109–123).
-
Rekombinante
menschliche Proteine, die in den vorstehend erwähnten Wirtsystemen exprimiert
werden, können
noch nicht-menschliche Glycoformen beinhalten (Raju et al., 2000
Annals Biochem. 283 (2): 123–132).
Im Besonderen kann dem Anteil der N-Glycane die terminale Sialinsäure fehlen,
die im Allgemeinen in menschlichen Glycoproteinen gefunden wird.
Wesentliche Anstrengungen waren auf die Entwicklung von Verfahren
zum Erhalt von Glycoproteinen ausgerichtet, die den menschlichen
Formen hinsichtlich der Struktur so nahe wie möglich sind oder die andere
therapeutische Vorteile besitzen. Glycoproteine, die spezifische
Glycoformen besitzen, können
besonders zweckdienlich sein, zum Beispiel für die zielgerichtete Lenkung
therapeutischer Proteine. So kann zum Beispiel das Anfügen von
einem oder mehreren Sialinsäure-Resten
an eine Glycan-Seitenkette die Lebensdauer eines therapeutischen
Glycoproteins nach Verabreichung in vivo erhöhen. Demgemäß können Säuger-Wirtzellen gentechnisch
verändert
sein, um den Umfang an terminalen Sialinsäuren in Glycoproteinen zu erhöhen, die
in den Zellen exprimiert sind. Alternativ dazu kann Sialinsäure vor der
Verabreichung in vitro an das Protein von Interesse konjugiert werden,
wobei eine Sialinsäuretransferase und
ein geeignetes Substrat verwendet werden. Zusätzlich wurden Veränderungen
in der Zusammensetzung des Wachstums-Mediums oder der Expression
von Enzymen, die an der menschlichen Glycosylierung beteiligt sind,
verwendet, um Glycoproteine herzustellen, die den menschlichen Formen ähnlicher
sind (S. Weikert, et al., Nature Biotechnology, 1999, 17, 1116–1121; Werner,
Noe, et al. 1998 Arzneimittelforschung 48 (8): 870–880; Weikert,
Papac et al., 1999; Andersen and Goochee 1994 Cur. Opin. Biotechnol.
5: 546–549;
Yang and Butler 2000 Biotechnol. Bioengin. 68 (4): 370–380). Alternativ
dazu können
gezüchtete
menschliche Zellen verwendet werden.
-
Alle
die bestehenden Systeme haben jedoch signifikante Nachteile. Nur
bestimmte therapeutische Proteine sind für die Expression in Säuger- oder
Pflanzen-Systemen
geeignet (z.B. solche, denen jegliche cytotoxische Wirkung fehlt
oder eine andere Wirkung, die für
Wachstum nachteilig ist). Tierische und Pflanzen-Zellkultur-Systeme sind gewöhnlich sehr
langsam, benötigen
häufig
mehr als eine Woche Wachstum unter sorgfältig kontrollierten Bedingungen,
um jede beliebige zweckmäßige Menge
des Proteins von Interesse herzustellen. Im Vergleich mit denjenigen
aus mikrobiellen Fermentations-Verfahren ist der Ertrag an Proteinen trotzdem
ungünstig.
Zusätzlich
benötigen
Zellkultur-Systeme im Allgemeinen komplexe und aufwendige Nährstoffe
und Co-Faktoren wie fötales
Kälberserum.
Darüber
hinaus kann das Wachstum durch programmierten Zelltod (Apoptose)
eingeschränkt
sein.
-
Darüber hinaus
sind tierische Zellen (insbesondere Säuger-Zellen) in hohem Maße empfänglich für virale
Infektion oder Kontamination. In einigen Fällen kann das Virus oder andere
infektiöse
Mittel das Wachstum der Kultur beeinträchtigen, während in anderen Fällen das
Mittel ein menschliches Pathogen sein kann, das das therapeutische
Protein-Produkt hinsichtlich seiner beabsichtigten Verwendung unbrauchbar
macht. Darüber
hinaus benötigen
zahlreiche Zellkultur-Verfahren die Verwendung von komplexen, temperatursensitiven, aus
Tieren stammenden Wachstumsmedium-Bestandteile, die Pathogene tragen
können,
wie bovine spongiforme Enzephalopathie (BSE)-Prionen. Solche Pathogene
sind schwierig nachzuweisen und/oder schwierig zu entfernen oder
zu sterilisieren, ohne das Wachstumsmedium zu beeinträchtigen.
In jedem Fall macht die Verwendung von tierischen Zellen zur Herstellung
therapeutischer Proteine kostspielige Qualitätskontrollen erforderlich,
um die Produkt-Sicherheit zu gewährleisten.
-
Zur
Herstellung großer
Volumina therapeutischer Proteine wie menschliches Serum-Albumin,
Gewebe-Plasminogen-Aktivator, monoclonale Antikörper, Hämoglobin, Kollagen, Fibrinogen
und andere können auch
transgene Tiere verwendet werden. Während transgene Ziegen und
andere transgene Tiere (Mäuse, Schafe,
Kühe, etc.)
gentechnisch verändert
werden können,
sodass sie therapeutische Proteine in hohen Konzentrationen in der
Milch produzieren, ist das Verfahren kostspielig, da jede Charge
sich einer drastischen Qualitätskontrolle
unterziehen muss. Tiere können
eine Vielzahl- tierischer oder menschlicher Pathogene tragen, einschließlich Bakterien,
Viren, Pilze und Prionen. Im Falle der Traberkrankheit und der bovinen
spongiformen Enzephalopathie kann das Testen, um eine Infektion
auszuschließen,
etwa ein Jahr dauern. Die Herstellung von therapeutischen Verbindungen
wird somit vorzugsweise in einer gut kontrollierten sterilen Umgebung
durchgeführt,
z.B. unter Bedingungen Guter Herstellerischer Praxis (GMP). Es ist
jedoch im Allgemeinen nicht durchführbar, Tiere in solchen Umgebungen
zu halten. Darüber
hinaus beruht transgene Tier-Technologie auf verschiedenen Tieren
und ist somit von Natur aus nicht einheitlich, wohingegen Zellen,
die in einem Fermenter wachsen, von einer gut charakterisierten
Original-Zellbank
(MCB) abstammen. Darüber
hinaus können
externe Faktoren wie unterschiedliche Nahrungsaufnahme, Krankheit
und mangelnde Homogenität
innerhalb einer Herde das Glycosylierungs-Muster des Endproduktes
beeinflussen. Es ist zum Beispiel in Menschen bekannt, dass verschiedene
Ernährungs-Angewohnheiten in
verschiedenen Glycosylierungs-Mustern resultieren.
-
Transgene
Pflanzen sind als eine potenzielle Quelle entwickelt worden, um
Proteine von therapeutischem Wert zu erhalten. Die Expression von
Proteinen in hohen Mengen in Pflanzen leidet jedoch unter dem Gene
Silencing, einem Mechanismus, durch den die Gene für stark
exprimierte Proteine in der nachfolgenden Pflanzen-Generation herunterreguliert
sind. Zusätzlich
fügen Pflanzen
Xylose und/oder α-1,3-gekoppelte
Fucose an N-Glycane von Proteinen an, was in Glycoproteinen resultiert,
die sich in der Struktur von tierischen unterscheiden und in Säugern immunogen
sind (Atlmann, Marz et al., 1995 Glycoconj. J. 12 (2); 150–155). Darüber hinaus
ist es im Allgemeinen nicht durchführbar Pflanzen in einer sterilen
oder GMP-Umgebung zu züchten
und die Gewinnung von Proteinen aus Pflanzen-Geweben ist kostspieliger als die Gewinnung
aus fermentierten Mikroorganismen. Glycoprotein-Herstellung, wobei
eukaryontische Mikroorganismen verwendet werden Der Mangel an einem
geeigneten Expressionssystem ist somit ein signifikantes Hindernis
für die
kostengünstige
und sichere Herstellung von rekombinanten menschlichen Glycoproteinen.
Die Herstellung von Glycoproteinen durch die Fermentation von Mikroorganismen
würde zahlreiche
Vorteile gegenüber
den bestehenden Systemen bieten. Auf Fermentation basierende Verfahren
können
zum Beispiel Folgendes bieten; (a) die schnelle Herstellung von
hohen Konzentrationen an Protein; (b) die Möglichkeit sterile, gut kontrollierte
Herstellungsbedingungen zu verwenden (z.B. GMP-Bedingungen); (c)
die Möglichkeit
einfache, chemisch definierte Wachstumsmedien zu verwenden; (d)
die Leichtigkeit genetischer Manipulation; (e) das Fehlen von kontaminierenden
menschlichen oder tierischen Pathogenen; (f) die Möglichkeit
eine große
Bandbreite an Proteinen zu exprimieren, einschließlich diejenigen,
die in Folge von Toxizität
etc. in der Zellkultur schlecht exprimiert werden; (g) die Leichtigkeit
der Protein-Gewinnung
(z.B. über
Sekretion in das Medium). Zusätzlich
sind Fermentations- Anlagen
im Allgemeinen wesentlich weniger kostspielig zu konstruieren als
Zellkultur-Anlagen.
-
Wie
vorstehend bemerkt, glycosylieren Bakterien, einschließlich Arten
wie Escherichia coli, die im Allgemeinen verwendet werden, um rekombinante
Proteine herzustellen, Proteine jedoch nicht in einer spezifischen
Art und Weise wie Eukaryonten. Verschiedene methylotrophe Hefen
wie Pichia pastoris, Pichia methanolica und Hansenula polymorpha
sind besonders zweckmäßig als
eukaryontische Expressionssysteme, da sie in der Lage sind zu hohen
Zelldichten zu wachsen und/oder große Mengen an rekombinantem
Protein zu sekretieren. Wie vorstehend bemerkt, unterscheiden sich
jedoch Glycoproteine, die in diesen eukaryontischen Mikroorganismen
exprimiert werden, wesentlich in der N-Glycan-Struktur von denjenigen in Tieren. Dies
hat die Verwendung von Hefe oder filamentösen Pilzen als Wirte für die Herstellung
von zahlreichen zweckmäßigen Glycoproteinen
verhindert.
-
Einige
Anstrengungen sind unternommen worden, um die Glycosylierungswege
von eukaryontischen Mikroorganismen zu modifizieren, um Glycoproteine
bereitzustellen, die für
die Verwendung als therapeutische Mittel in Säugern besser geeignet sind.
Es sind zum Beispiel einige Glycosyltransferasen vereinzelt cloniert und
in S. cerevisiae (GalT, GnT I), Aspergillus nidulans (GnT I) und
anderen Pilzen (Yoshida et al., 1999, Kalsner et al., 1995 Glycoconj.
J. 12(3): 360–370,
Schwientek et al., 1995) cloniert worden. Es wurden jedoch keine N-Glycane
mit menschlichen Eigenschaften erhalten.
-
Hefen
produzieren eine Vielzahl von Mannosyltransferasen, z.B. 1,3-Mannosyltransferasen
(z.B. MNN1 in S. cerevisiae) (Graham and Emr, 1991 J. Cell Biol.
114 (2): 207–218),
1,2-Mannosyltransferasen (z.B. die KTR/KRE-Familie von S. cerevisiae),
1,6-Mannosyltransferasen (OCH1 von S. cerevisiae), Mannosylphosphat-Transferasen
(MNN4 und MNN6 von S. cerevisiae) und zusätzliche Enzyme, die an endogenen
Glycosylierungs-Reaktionen beteiligt sind. Viele dieser Gene sind
vereinzelt deletiert worden, wobei lebensfähige Organismen entstanden
sind, die veränderte
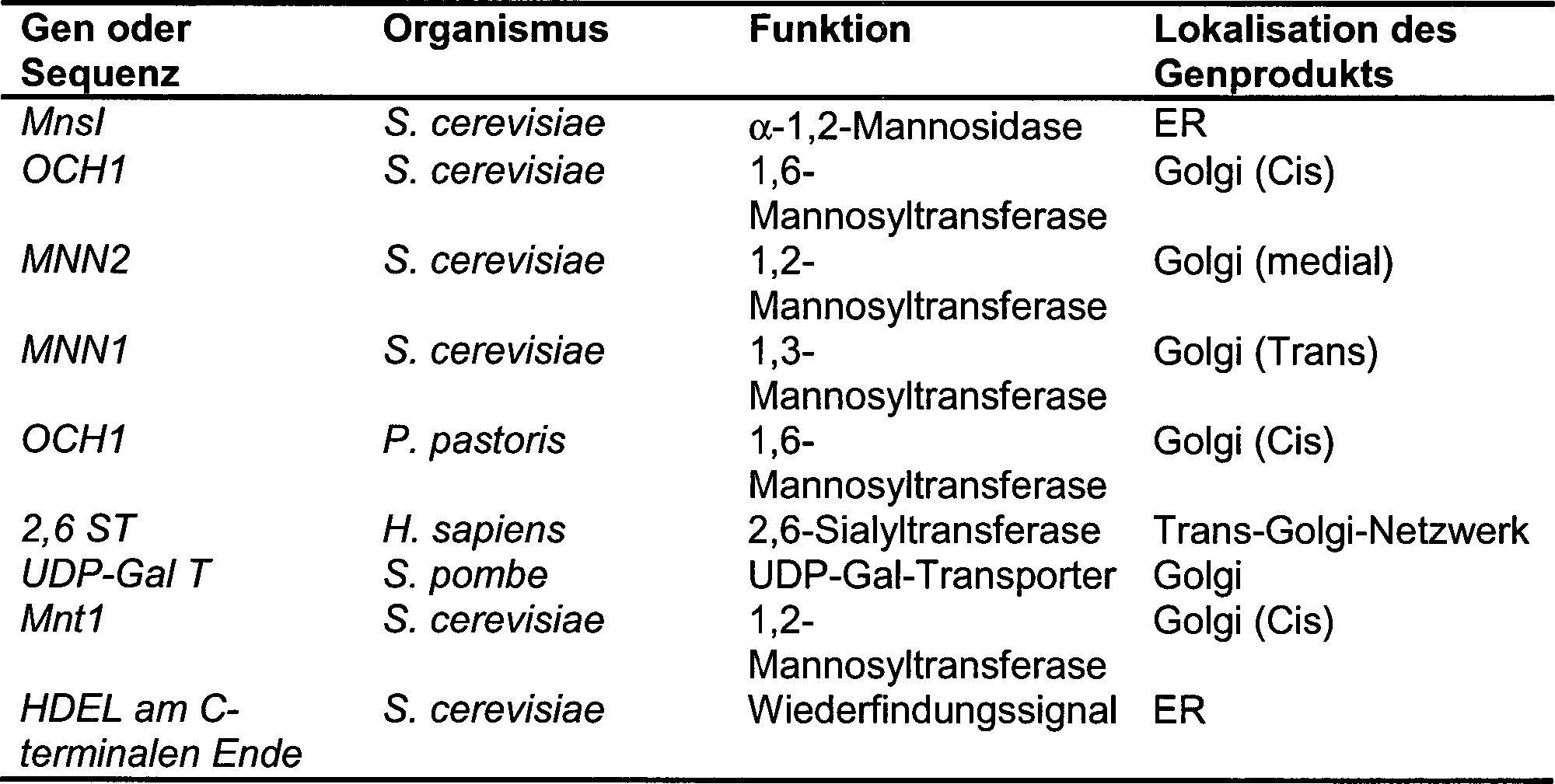
Glycosylierungs-Profile besitzen. Beispiele sind in Tabelle 1 gezeigt.
-
Tabelle
1. Beispiele von Hefe-Stämmen,
die eine veränderte
Mannosylierung besitzen
-
Zusätzlich offenbart
die Japanische Patent-Anmeldung mit der Veröffentlichungs-Nr. 8-336387
einen mutierten OCH1-Stamm von Pichia pastoris. Das OCH1-Gen codiert
1,6-Mannosyltransferase, die eine Mannose an die Glycan-Struktur Man8GlcNAc2 anfügt, wobei
Man9GlcNAc2 erhalten
wird. Die Man9GlcNAc2-Struktur ist dann
ein Substrat für
weitere Mannosylierung in vivo, was dann zu den hypermannosylierten
Glycoproteinen führt,
die für
Hefen charakteristisch sind und im Allgemeinen mindestens 30 bis
40 Mannose-Reste pro N-Glycan haben können. In dem mutierten OCH1-Stamm
sind Proteine angereichert, die mit Man8GlcNAc2 glycosyliert sind, und Hypermannosylierung
tritt nicht auf. Die Struktur Man8GlcNAc2 ist jedoch kein Substrat für tierische
Glycosylierungs-Enzyme wie menschliche UDP-GlcNAc-Transferase I und entsprechend
ist das Verfahren für
die Herstellung von Proteinen mit menschlichen Glycosylierungs-Mustern
nicht zweckmäßig.
-
Martinet
und Mitarbeiter (Biotechnol. Lett. 1998, 20 (12), 1171–1177) berichteten über die
Expression einer α-1,2-Mannosidase
von Trichoderma reesei in P. pastoris. Es wurde einiges Mannosetrimmen
der N-Glycane eines Modell-Proteins beobachtet. Das Modell-Protein
hatte jedoch keine N-Glycane mit der Struktur Man5GlcNAc2, was als ein Zwischenprodukt für die Erzeugung
von komplexen N-Glycanen
notwendig wäre. Demgemäß ist das
Verfahren nicht zweckmäßig für die Herstellung
von Proteinen mit menschlichen oder tierischen Glycosylierungs-Mustern.
-
In ähnlicher
Weise exprimierten Chiba und Mitarbeiter 1998 α-1,2-Mannosidase von Aspergillus saitoi in
der Hefe Saccharomyces cerevisiae. Eine Signalpeptid-Sequenz (His-Asp-Glu-Leu)
wurde gentechnisch in die exogene Mannosidase rekombiniert, um das
Zurückhalten
in dem endoplasmatischen Retikulum zu verstärken. Zusätzlich handelte es sich bei
dem Hefe-Wirt um eine Mutante, der drei Enzym-Aktivitäten fehlten, die
mit Hypermannosylierung von Proteinen assoziiert sind: 1,6-Mannosyltransferase
(OCH1); 1,3-Mannosyltransferase (MNN1); und Mannosylphosphat-Transferase
(MNN4). Die N-Glycane des dreifach mutierten Wirtes bestanden somit
aus der Struktur Man8GlcNAc2,
die gegenüber
den High-Mannose-Formen bevorzugt wird, die in Wildtyp-S. cerevisiae
gefunden werden. In Anwesenheit der stark exprimierten gentechnisch
veränderten
Mannosidase wurden die N-Glycane eines Modellproteins (Carboxypeptidase
Y) ausgetrimmt, wobei ein Gemisch, bestehend aus 27 Mol% Man5GlcNAc2, 22 Mol%
Man6GlcNAc2, 22
Mol% Man7GlcNAc2 und
29 Mol% Man8GlcNAc2 erhalten
wurde. Das Trimmen der endogenen Zellwand-Glycoproteine war weniger
effizient, nur 10 Mol% der N-Glycane hatten die gewünschte Man5GlcNAc2-Struktur. Über Chibas
Arbeit ist auch in EP-A1 1211311 berichtet worden. Obwohl Chiba
eine mehr als ausreichende Menge seiner Mannosidase in hoher Kopienzahl
exprimierte und er relativ geringfügige Erträge erhielt, schlägt er vor,
dass sogar noch mehr Enzym exprimiert werden sollte, um das Problem
der niedrigen Erträge
auszuräumen.
Er schlug niemals vor, dass das gewünschte Enzym so ausgewählt werden
sollte, dass es in der ausgewählten
Wirtszelle an dem subzellulären
Ort, an dem es sich befindet, optimal arbeitet, um sein Substrat
umzuwandeln.
-
Da
ausschließlich
die Man5GlcNAc2-Glycane
für eine
weitere enzymatische Umwandlung zu menschlichen Glycoformen zugänglich sind,
ist das Verfahren nicht wirksam für die Herstellung von Proteinen,
die menschliche Glycosylierungs-Muster besitzen. In Proteinen, die
eine einzige N-Glycosylierungsstelle besitzen, hätten mindestens 73 Mol% eine
unkorrekte Struktur. In Proteinen, die zwei bzw. drei N-Glycosylierungsstellen besitzen,
hätten
mindestens 93 bzw. 98 Mol% eine unkorrekte Struktur. Solche niedrigen
Wirkungsgrade bezüglich
der Umwandlung sind nicht befriedigend für die Herstellung von therapeutischen
Mitteln, insbesondere da die Abtrennung von Proteinen, die verschiedene
Glycoformen besitzen, im Allgemeinen kostspielig und schwierig ist.
-
Mit
dem Ziel, ein mehr menschenähnliches
Glycoprotein bereitzustellen, das von einem Pilz als Wirt abstammt,
offenbart das US-Patent Nr. 5,834,251 von Maras und Contreras ein
Verfahren zur Herstellung eines Hybrid-Glycoproteins, das von Trichoderma
reesei abstammt. Ein Hybrid-N-Glycan hat an dem Man-α1-6-Arm des
Kerns ausschließlich
Mannose-Reste und eine oder zwei komplexe Antennen an dem Man-α1-3-Arm.
Während
diese Struktur Nutzen hat, hat das Verfahren den Nachteil, dass
zahlreiche enzymatische Schritte in vitro durchgeführt werden
müssen,
was kostspielig und zeitraubend ist. Isolierte Enzyme sind teuer
herzustellen und zu erhalten, können
ungewöhnliche
und kostspielige Substrate benötigen
(z.B. UDP-GlcNAc) und neigen unter den verwendeten Bedingungen zu
Verlust an Aktivität
und/oder zu Proteolyse.
-
Deshalb
ist es ein Ziel der vorliegenden Erfindung ein System und Verfahren
für humanisierende
Glycosylierung von rekombinanten Glycoproteinen bereitzustellen,
die in Pichia pastoris und anderen niederen Eukaryonten wie Hansenula
polymorpha, Pichia stiptis, Pichia methanolica, Pichia sp, Kluyveromyces
sp, Candida albicans, Aspergillus nidulans und Trichoderma reesei
exprimiert werden.
-
ZUSAMMENFASSUNG DER ERFINDUNG
-
Es
wurden Zelllinien entwickelt, die gentechnisch modifizierte Glycosylierungswege
besitzen, die es ihnen erlauben eine Folge von enzymatischen Reaktionen
durchzuführen,
die die Prozessierung von Glycoproteinen in Menschen nachahmen.
Rekombinante Proteine, die in diesen gentechnisch veränderten
Wirten exprimiert werden, stellen Glycoproteine bereit, die ihren
menschlichen Gegenstücken ähnlicher,
wenn nicht im Wesentlichen mit diesen identisch sind. Die niederen
Eukaryonten, die im Allgemeinen High-Mannose-enthaltende N-Glycane
produzieren, einschließlich
unizelluläre
und multizelluläre
Pilze wie Pichia pastoris, Hansenula polymorpha, Pichia stiptis,
Pichia methanolica, Pichia sp., Klyveromyces sp., Candida albicans,
Aspergillus nidulans und Trichoderma reesei werden modifiziert,
sodass sie N-Glycane wie Man5GlcNAc2 oder andere Strukturen entlang des menschlichen
Glycosylierungswegs produzieren. Dies wird erreicht, indem eine Kombination
von gentechnischen Verfahren und/oder einer Auswahl von Stämmen verwendet
wird, die: keine bestimmten Enzyme exprimieren, die die unerwünschten komplexen
Strukturen erzeugen, die für
die Glycoproteine von Pilzen charakteristisch sind, die exogene
Enzyme exprimieren, die so ausgewählt wurden, dass sie entweder
unter den Bedingungen, die in dem Pilz vorliegen, wo Aktivität erwünscht ist,
optimale Aktivität besitzen
oder die auf ein Organell gerichtet sind, wo optimale Aktivität erreicht
wird, und Kombinationen daraus, in denen der genetisch veränderte Eukaryont
multiple exogene Enzyme exprimiert, die erforderlich sind, um "Menschen-ähnliche"-Glycoproteine zu produzieren.
-
In
einer ersten Ausführungsform
betrifft die Erfindung eine Wirtszelle, die ein unizellulärer oder
filamentöser
Pilz ist, der keine α-1,6-Mannosyltransferase-Aktivität, bezogen
auf das N-Glycan auf einem Glycoprotein, aufweist, und die in ihrem
endoplasmatischen Retikulum (ER) oder Golgi-Apparat ein Hybridenzym hat,
das ausgewählt
ist, um optimale Aktivität
im ER oder Golgi der Wirtszelle zu haben, sodass die Wirtszelle 50
bis 100 Mol% Man5GlcNAc2 auf
einem Substrat-Glycoprotein
bilden kann, wobei das Hybridenzym umfasst:
- (a)
eine exogene Mannosidase-katalytische Domäne mit optimaler Aktivität im ER
oder Golgi bei einem pH-Wert zwischen 5,1 oder 8,0; verbunden mit
- (b) einem zellulären
Zielsignal-Peptid, das normalerweise nicht mit der katalytischen
Domäne
von (a) verbunden ist, wobei das zelluläre Zielsignal-Peptid die exogene
Mannosidase-katalytische Domäne
in das ER oder den Golgi-Apparat bringt.
-
Somit
betrifft die Erfindung einen Mikroorganismus, der gentechnisch verändert ist,
um ein exogenes α-1,2-Mannosidase-Enzym
zu exprimieren, das einen optimalen pH-Wert zwischen 5,1 und 8,0
besitzt, vorzugsweise zwischen 5,9 und 7,5. Somit ist das exogene
Enzym auf das endoplasmatische Retikulum oder den Golgi-Apparat des Wirtsorganismus
gerichtet, wo es N-Glycane wie Man8GlcNAc2 derart trimmt, dass Man5GlcNAc2 erhalten wird. Die letztere Struktur ist
zweckmäßig, weil
sie mit einer Struktur übereinstimmt,
die in Säugern,
insbesondere Menschen, gebildet wird; sie ist ein Substrat für weitere
Glycosylierungs-Reaktionen in vivo und/oder in vitro, die ein endgültiges N-Glycan
produzieren, das dem ähnlich
oder zu dem identisch ist, das in Säugern, insbesondere in Menschen
gebildet wird; und sie ist kein Substrat für Hypermannosylierungs-Reaktionen,
die in vivo in Hefe und anderen Mikroorganismen auftreten und die
ein Glycoprotein hochgradig immunogen in Säugern machen.
-
In
einer zweiten Ausführungsform
betrifft die Erfindung ein Verfahren zur Herstellung der Wirtszelle, die
hierin offenbart ist, wobei das Verfahren den Schritt umfasst, diesen
unizellulären
oder filamentösen
Pilz mit einem Nucleinsäure-Molekül zu transformieren,
das ein Hybridenzym codiert, das ausgewählt ist, um optimale Aktivität im ER
oder Golgi der Wirtszelle zu haben, sodass die Wirtszelle 50 bis
100 Mol% Man5GlcNAc2 auf
einem Substrat-Glycoprotein bilden kann, wobei das Hybridenzym umfasst:
- (a) eine exogene Mannosidase-katalytische Domäne mit optimaler
Aktivität
im ER oder Golgi bei einem pH-Wert zwischen 5,1 und 8,0; verbunden
mit
- (b) einem zellulären
Zielsignal-Peptid, das normalerweise nicht mit der katalytischen
Domäne
von (a) verbunden ist, wobei das zelluläre Ziesignal-Peptid die exogene
katalytische Domäne
in das ER oder den Golgi-Apparat bringt.
-
In
einer dritten Ausführungsform
betrifft die Erfindung ein Verfahren zur Herstellung eines Glycoproteins
in einer Wirtszelle, die ein unizellulärer oder filamentöser Pilz
ist, der keine α-1,6-Mannosyltransferase-Aktivität in Bezug
auf das N-Glycan auf einem Glycoprotein aufweist, wobei das Verfahren
die Schritte umfasst:
- (a) Exprimieren des Glycoproteins
in einer Population von Wirtszellen, die mit einer DNA-Bibliothek
transformiert sind, die mindestens zwei verschiedene genetische
Konstrukte umfasst, wobei mindestens eines davon ein DNA-Fragment
umfasst, das ein Hybridenzym codiert, umfassend:
- (i) eine Glycosylierungsenzym-katalytische Domäne mit optimaler
Aktivität
im ER oder Golgi bei einem pH-Wert zwischen 5,1 und 8,0; verbunden
mit
- (ii) einem zellulären
Zielsignal-Peptid, das normalerweise nicht mit der katalytischen
Domäne
von (i) verbunden ist, wobei das zelluläre Zielsignal-Peptid auf das
ER oder den Golgi-Apparat der Wirtszelle abzielt; und
- (b) Auswählen
der Wirtszelle, die ein Glycoprotein mit einem gewünschten
Glycosylierungs-Muster auf dem Glycoprotein zeigt.
-
Somit
ist der Glycosylierungs-Signalweg eines eukaryontischen Mikroorganismus
in dieser Ausführungsform
modifziert, indem (a) eine DNA-Bibliothek
erzeugt wird, die mindestens zwei Gene beinhaltet, die exogene Glycosylierungs-Enzyme
codieren; (b) indem der Mikroorganismus mit der Bibliothek transformiert wird,
um eine genetisch gemischte Population zu erzeugen, die mindestens
zwei voneinander verschiedene exogene Glycosylierungs-Enzyme exprimiert;
(c) indem aus der Population ein Mikroorganismus ausgewählt wird,
der den gewünschten
Glycosylierungs-Phänotyp
besitzt. In einer anderen Ausführungsform
beinhaltet eine DNA-Bibliothek chimäre Gene, wobei jedes eine Protein-Lokalisierungs-Sequenz
und eine katalytische Aktivität
codiert, die mit Glycosylierung in Beziehung steht. Entsprechend
stellt die Erfindung in einer vierten Ausführungsform eine DNA-Bibliothek
bereit, die genetische Konstrukte umfasst, wobei die Konstrukte
ein DNA-Fragment umfassen, das ein zelluläres Zielsignal-Peptid codiert, das
auf das endoplasmatische Retikulum oder den Golgi-Apparat abzielt,
das im Leserahmen mit einem DNA-Fragment ligiert ist, das ein Glycosylierungs-Enzym
oder ein katalytisch aktives Fragment davon codiert, das in dem
ER oder Golgi bei einem pH-Wert zwischen 5,1 und 8,0 optimale Aktivität besitzt,
wobei die DNA-Bibliothek mindestens zwei DNA-Fragmente umfasst,
die ein zelluläres
Zielsignal-Peptid codieren, und mindestens zwei DNA-Fragmente umfasst, die
exogene Glycosylierungs-Enzyme codieren. Organismen, die unter Verwendung
dieses Verfahrens modifiziert wurden, sind zweckmäßig zur
Herstellung von Glycoproteinen, die ein Glycosylierungs-Muster aufweisen,
das dem von Säugern,
insbesondere Menschen, ähnlich
oder dazu identisch ist.
-
In
einer fünften
Ausführungsform
ist der Glycosylierungs-Signalweg modifiziert, sodass er ein Zucker-Nucleotid-Transport-Enzym
exprimiert. In einer bevorzugten Ausführungsform wird auch ein Nucleotid-Diphosphatase-Enzym
exprimiert. Der Transporter und die Diphosphatase verbessern die
Wirksamkeit der gentechnisch veränderten
Glycosylierungsschritte, indem die geeigneten Substrate für die Glycosylierungs-Enzyme
in den geeigneten Kompartimenten bereitgestellt werden, die kompetitive
Produkthemmung reduziert wird und das Entfernen von Nucleosid-Diphosphaten
verstärkt
wird.
-
Schließlich betrifft
die Erfindung eine Glycoprotein-Zusammensetzung, die durch die hierin
offenbarten Verfahren erhältlich
ist und die 50 bis 100 Mol% von Man5GlcNAc2 umfasst, das in vivo durch GnTI zu GlcNAcMan5GlcNAc2 umgewandelt
wird, wobei diesem Glycoprotein Galactose, Fucose und terminale
Sialinsäure
fehlen.
-
BESCHREIBUNG DER FIGUREN
-
1A ist
ein schematisches Diagramm des typischen Pilz-N-Glycosylierungs-Signalwegs.
-
1B ist
ein schematisches Diagramm eines typischen menschlichen N-Glycosylierungs-Signalwegs.
-
AUSFÜHRLICHE BESCHREIBUNG DER ERFINDUNG
-
Die
Verfahren und rekombinanten niederen Eukaryonten-Stämme, die
hierin beschrieben sind, werden verwendet, um "humanisierte Glycoproteine" herzustellen. Die
rekombinanten niederen Eukaryonten werden hergestellt, indem niedere
Eukaryonten, die eines oder mehrere Enzyme, die bei der Herstellung
von High-Mannose-Strukturen
beteiligt sind, nicht exprimieren, derart gentechnisch verändert werden,
dass sie die Enzyme exprimieren, die erforderlich sind, um Menschenähnliche
Zucker zu produzieren. Wie hierin verwendet, ist ein niederer Eukaryont
ein einzelliger oder filamentöser
Pilz. Wie hierin verwendet, bezieht sich ein "humanisiertes Glycoprotein" auf ein Protein,
an das N-Glycane angeheftet sind, die weniger als vier Mannose-Reste
beinhalten, und die synthetischen Zwischenprodukte (die ebenfalls
zweckmäßig sind
und in vitro weiter manipuliert werden können), die mindestens fünf Mannose-Reste
besitzen. In einer bevorzugten Ausführungsform enthalten die Glycoprotein,
die in dem rekombinanten niederen eukaryontischen Stamm produziert werden,
mindestens 27 Mol% des Man5-Zwischenprodukts.
Dies wird erreicht, indem eine bessere Mannosidase cloniert wird,
d.h. ein Enzym, das so ausgewählt
wird, dass es unter den Bedingungen, die in dem Organismus vorliegen,
an der Stelle, an der die Proteine glycosyliert werden, optimale
Aktivität
besitzt oder durch Lenken des Enzyms zu dem Organell, wo die Aktivität gewünscht ist.
-
In
einer bevorzugten Ausführungsform
werden eukaryontische Stämme
verwendet, die ein oder mehrere Enzyme nicht exprimieren, die an
der Produktion von High-Mannose-Strukturen beteiligt sind. Diese Stämme können durch
gentechnische Manipulation erzeugt werden oder einer von diesen
zahlreichen Mutanten sein, die bereits in Hefen beschrieben sind,
einschließlich
einer Hypermannosylierungs-minus(OCH1)-Mutante in Pichia pastoris.
-
Die
Stämme
können
für jeweils
ein einzelnes Enzym gentechnisch verändert werden oder es kann eine
Bibliothek von Genen erzeugt werden, die potenziell zweckmäßige Enzyme
codiert, und diejenigen Stämme
können
ausgewählt
werden, die Enzyme mit optimalen Aktivitäten besitzen oder die die am
meisten "Menschenähnlichen" Glycoproteine produzieren.
-
Niedere
Eukaryonten, die in der Lage sind Glycoproteine herzustellen, die
das angeheftete N-Glycan Man5GlcNAc2 besitzen, sind besonders zweckmäßig, denn,
da (a) ihnen eine hochgradige Mannosylierung fehlt (z.B. mehr als
acht Mannosen pro N-Glycan oder im Besonderen 30–40 Mannosen), zeigen sie reduzierte Immunogenität in Menschen;
und (b) das N-Glycan ist ein Substrat für weitere Glycosylierungs-Reaktionen, wobei
eine sogar noch mehr Menschen-ähnliche
Glycoform gebildet wird, z.B. durch die Wirkung von GlcNAc-Transferase
I, wobei GlcNAcMan5GlcNAc2 gebildet
wird. Man5GlcNAc2 muss
in vivo in einem hohen Ertrag gebildet werden, zumindest vorübergehend,
da alle nachfolgenden Glycosylierungs-Reaktionen Man5GlcNAc2 oder ein Derivat davon benötigen. Entsprechend
wird ein Ertrag von mehr als 27 Mol%, mehr bevorzugt ein Ertrag
von 50–100
Mol% Glycoproteine erhalten, in denen ein hoher Anteil von N-Glycanen Man5GlcNAc2 besitzen.
Bevorzugte erfindungsgemäße Wirtszellen
erbringen 50–100
Mol% Glycoproteine, in denen N-Glycane Man5GlcNAc2 besitzen, das in vivo durch GlcNAc-Transferase
I umgewandelt wurde. Es ist dann möglich in vitro weitere Glycosylierungs-Reaktionen
durchzuführen,
wobei zum Beispiel das Verfahren des US-Patents Nr. 5,834,251 von
Maras und Contreras verwendet wird. In einer bevorzugten Ausführungsform
wird mindestens eine weitere Glycosylierungs-Reaktion in vivo durchgeführt. In
einer hochgradig bevorzugten Ausführungsform davon werden aktive
Formen weiterer Glycosylierungs-Enzyme in dem endoplasmatischen
Retikulum und/oder dem Golgi-Apparat exprimiert.
-
Wirts-Mikroorganismen
-
Hefe
und filamentöse
Pilze sind beide für
die Herstellung von rekombinanten Proteinen, sowohl intrazelluläre als auch
sekretierte, erfolgreich verwendet worden (Cereghino, J. L. and
J. M. Cregg 2000 FEMS Microbiology Reviews 24 (1): 45–66; Harkki,
A., et al. 1989 Bio-Technology 7 (6): 596; Berka, R. M., et al.
1992 Abstr. Papers Amer. Chem. Soc. 203: 121-BIOT; Svetina, M.,
et al. 2000 J. Biotechnol. 76 (2–3): 245–251.
-
Obwohl
die Glycosylierung in Hefe und Pilzen sich von der in Menschen stark
unterscheidet, werden einige Elemente gemeinsam benutzt. Der erste
Schritt, der Transfer der Kern-Oligosaccharid-Struktur an das nascierende
Protein ist in allen Eukaryonten, einschließlich Hefen, Pilze, Pflanzen
und Menschen, hoch konserviert (vergleiche 1A und 1B).
Die nachfolgende Prozessierung des Kern-Oligosaccharids unterscheidet sich jedoch
signifikant in Hefe und beinhaltet das Anhängen von einigen Mannose-Zuckern.
Dieser Schritt wird durch Mannosyltransferasen katalysiert, die
sich in dem Golgi befinden (z.B. OCH1, MNT1, MNN1, etc.), die sequenziell
Mannose-Zucker an das Kern-Oligosaccharid anheften. Die resultierende
Struktur ist für die
Herstellung Menschen-ähnlicher
Proteine unerwünscht
und somit ist es wünschenswert,
die Mannosyltransferase-Aktivität zu reduzieren
oder auszuschalten. Für
Mutanten von S. cerevisiae, die für Mannosyltransferase-Aktivität defizient
sind (z.B. och1- oder mnn9-Mutanten), konnte gezeigt werden, dass
sie nicht letal sind und in dem Oligosaccharid von Hefe-Glycoproteinen einen
reduzierten Mannose-Gehalt zeigen. In Abhängigkeit von dem jeweiligen
endogenen Glycosylierungs-Muster des Wirts kann es auch erforderlich
sein, dass andere Oligosaccharid-prozessierende Enzyme wie Mannosylphosphat-Transferase ebenfalls
ausgeschaltet werden müssen.
Nach der Reduktion unerwünschter
endogener Glycosylierungs-Reaktionen muss die Bildung von komplexen
N-Glycanen das Wirtssystem eingebracht werden. Dies erfordert die
stabile Expression von einigen Enzymen und Zucker-Nucleotid-Transportern.
Darüber
hinaus müssen
diese Enzyme auf eine Art und Weise lokalisiert werden, dass eine
sequenzielle Prozessierung der reifenden Glycosylierungs-Struktur gewährleistet
ist.
-
Ziel-Glycoproteine
-
Die
hierin beschriebenen Verfahren sind für die Herstellung von Glycoproteinen
zweckmäßig, insbesondere
Glycoproteine, die therapeutisch in Menschen verwendet werden. Solche
therapeutischen Proteine werden typischerweise durch Injektion,
oral, pulmonal oder auf andere Art und Weise verabreicht.
-
Beispiele
für geeingete
Ziel-Glycoproteine beinhalten, ohne Einschränkung: Erythropoietin, Cytokine wie
Interferon-α,
Interferon-β,
Interferon-γ,
Interferon-ω und
Granulocyten-CSF, Koagulationsfaktoren wie Faktor VIII, Faktor IX
und menschliches Protein C, lösliche
IgE-Rezeptor-α-Kette,
IgG, IgM, Urokinase, Chymase und Harnstoff-Trypsin-Inhibitor, IGF-bindendes
Protein, epidermaler Wachstumsfaktor, Wachstumshormon-feisetzender
Faktor, Annexin V-Fusionsprotein,
Angiostatin, vaskulärer
endothelialer Wachstumsfaktor-2, myeloider Vorläufer-Inhibitor-Faktor-1 und
Osteoprotegerin.
-
Verfahren zur Herstellung
von Glycoproteinen, die das N-Glycan Man5GlcNAc2 umfassen
-
Der
erste Schritt beinhaltet die Auswahl oder die Erzeugung eines niederen
Eukaryonten, der in der Lage ist eine spezifische Vorläufer-Struktur
von Man5GlcNAc2 zu
produzieren, die in der Lage ist in vivo durch die Wirkung einer
GlcNAc-Transferase I GlcNAc aufzunehmen. Dieser Schritt erfordert
die Bildung einer bestimmten isomeren Struktur von Man5GlcNAc2. Diese Struktur muss innerhalb der Zelle
in einem hohen Ertrag gebildet werden (mehr als 30%), da alle nachfolgenden
Manipulationen von der Anwesenheit dieser Vorstufe abhängig sind.
Man5GlcNAc2-Strukturen
sind für
die Bildung von komplexem N-Glycan erforderlich, ihr Vorliegen ist
jedoch in keinster Weise ausreichend, da Man5GlcNAc2 in verschiedenen isomeren Formen auftreten kann,
die als ein Substrat für
GlcNAc-Transferase
I dienen können
oder nicht. Die meisten Glycosylierungs-Reaktionen sind nicht vollständig und
somit enthält
ein bestimmtes Protein im Allgemeinen eine Auswahl unterschiedlicher
Kohlenhydrat-Strukturen (d.h. Glycoformen) auf seiner Oberfläche. Die
bloße
Anwesenheit von Spuren-Mengen (weniger als 5%) einer bestimmten
Struktur wie Man5GlcNAc2,
ist von geringer praktischer Relevanz. Es ist die Bildung eines
bestimmten GLcNAc-Transferase I akzeptierenden Zwischenprodukts (Struktur
I) in hohem Ertrag (oberhalb von 30%), die erforderlich ist. Die
Bildung dieses Zwischenprodukts ist notwendig und erlaubt im Nachfolgenden
die in vivo-Synthese von komplexen N-Glycanen.
-
Um
die Struktur in vivo bereitzustellen, kann man solche niederen Eukaryonten
aus der Natur auswählen
oder alternativ dazu vorliegende Pilze oder andere niedere Eukaryonten
gentechnisch manipulieren. Es wurde bisher kein niederer Eukaryont
gezeigt, der solche Strukturen in vivo in höherem Maße als 1,8% in Bezug auf die
gesamten N-Glycane bereitstellt (Mayas et al., 1997), sodass ein
gentechnisch manipulierter Organismus bevorzugt wird. Verfahren
wie diejenigen, die im US-Patent Nr. 5,595,900 beschrieben sind,
können verwendet
werden, um die Abwesenheit oder Anwesenheit von bestimmten Glycosyltransferasen,
Mannosidasen und Zucker-Nucleotid-Transportern in einem Zielorganismus
von Interesse zu identifizieren.
-
Inaktivierung von Pilz-Glycosylierungs-Enzymen
wie 1,6-Mannosyltransferase
-
Das
hierin beschriebene Verfahren kann verwendet werden, um das Glycosylierungs-Muster
einer großen
Bandbreite niederer Eukaryonten gentechnisch zu manipulieren (z.B.
Hansenula polymorpha, Pichia stiptis, Pichia methanolica, Pichia
sp, Kluyveromyces sp, Candida albicans, Aspergillus nidulans, Trichoderma reesei
etc.). Pichia pastoris wird verwendet, um die erforderlichen Manipulationsschritte
beispielhaft darzustellen. Ähnlich
wie andere niedere Eukaryonten prozessiert P. pastoris Man>9GlcNAc2-Strukturen in dem ER mit einer 1,2-α-Mannosidase,
wobei Man8GlcNAc2 erhalten
wird. Durch die Wirkung einiger Mannosyltransferasen wird diese
Struktur dann in hypermannosylierte Strukturen umgewandelt (Man9GlcNAc2), die auch
als Mannane bekannt sind. Zusätzlich
wurde gefunden, dass P. pastoris in der Lage ist nicht-terminale
Phophatgruppen über
die Wirkung von Mannosylphosphat-Transferasen an die Kohlenhydrat-Struktur
anzuhängen.
Das ist im Gegensatz zu den Umsetzungen, die in Säugerzellen
gefunden werden, die das Entfernen von Mannose-Zucker beinhalten,
im Gegensatz zu deren Hinzufügung.
Es ist von besonderer Wichtigkeit die Fähigkeit des Pilzes auszuschalten,
die vorliegende Man8GlcNAc2-Struktur
zu hypermannolysieren. Das kann erreicht werden, indem entweder
ein Pilz ausgewählt
wird, der nicht hypermannosyliert ist oder indem ein solcher Pilz
gentechnisch manipuliert wird.
-
Gene,
die an diesem Prozess beteiligt sind, wurden in Pichia pastoris
identifiziert und durch Erzeugen von Mutationen in diesen Genen
ist man in der Lage die Herstellung von "unerwünschten" Glycoformen zu reduzieren. Solche Gene
können
durch Homologie zu bestehenden Mannosyltransferasen (z.B. OCH1,
MNN4, MNN6, MNN1) identifiziert werden, die in anderen niederen
Eukaryonten wie C. albicans, Pichia angusta oder S. cerevisiae gefunden
werden, oder indem der Wirtsstamm mutiert wird und auf einen Phänotyp mit
reduzierter Mannosylierung selektiert wird. Basierend auf Homologien
zwischen bekannten Mannosyltransferasen und Mannosylphosphat-Transferasen
kann man entweder PCR-Primer entwerfen, wofür Beispiele in Tabelle 2 gezeigt
sind, oder Gene oder Genfragmente, die solche Enzyme codieren, als
Sonden verwenden, um Homologe in den DNA-Bibliotheken der Ziel-Organismen
zu identifizieren. Alternativ dazu kann es möglich sein bestimmte Phänotypen
in verwandten Organismen zu ergänzen.
Um zum Beispiel das Gen oder die Gene zu erhalten, die 1,6-Mannosyltransferase-Aktivität in P.
pastoris codieren, würde
man die nachfolgenden Schritte durchführen. OCH1-Mutanten von S.
cerevisiae sind temperatursensitiv und wachsen langsam bei erhöhten Temperaturen.
Somit kann man funktionelle Homologe von OCH1 in P. pastoris identifizieren,
indem man eine OCH1-Mutante von S. cerevisiae mit einer DNA- oder
cDNA-Bibliothek aus P. pastoris ergänzt. Solche Mutanten von S.
cerevisiea können
auf der Seite http://genome-www.stanford.edu/Saccharomyces/ gefunden
werden und sind auf der Seite http://www.resgen.com/products/YEASTD.php3
kommerziell erhältlich.
Mutanten, die bei erhöhter
Temperatur einen Phänotyp
mit normalem Wachstum zeigen, nachdem sie mit einer P. pastoris-DNA-Bibliothek
transformiert worden waren, tragen wahrscheinlich ein OCH1-Homologes
von P. pastoris. Eine solche Bibliothek kann erzeugt werden, indem
chromosomale DNA von P. pastoris mit einem geeigneten Restriktionsenzym
partiell gespalten wird und die gespaltene DNA nach Inaktivierung
des Restriktionsenzyms in einen geeigneten Vektor ligiert wird,
der mit einem passenden Restriktionsenzym gespalten worden war.
Geeignete Vektoren sind pRS314, ein Plasmid mit niedriger Kopienzahl
(CEN6/ARS4), das auf pBluescript basiert, das den Trp1-Marker enthält (Sikorski,
R. S., and Hieter, P., 1989, Genetics 122, S. 19–27), oder pFL44S, ein Plasmid
von hoher Kopienzahl (2μ),
das auf einem modifizierten pUC19 basiert, das den URA3-Marker enthält (Bonneaud,
N., et al., 1991, Yeast 7, S. 609–615). Solche Vektoren werden
gewöhnlich
von akademischen Wissenschaftlern verwendet oder ähnliche
Vektoren sind von einer Vielzahl unterschiedlicher Anbieter erhältlich,
wie Invitrogen (Carlsbad, CA), Pharmacia (Piscataway, NJ), New England
Biolabs (Beverly, MA). Beispiele sind pYES/GS, 2μ-Replikationsursprung, basierend auf
dem Hefe-Expressionsplasmid von Invitrogen, oder Yep24, ein Clonierungs-Vehikel
von New England Biolabs. Nach Ligierung der chromosomalen DNA und
des Vektors kann man die DNA-Bibliothek in einen Stamm von S. cerevisiae
mit einer spezifischen Mutation transformieren und hinsichtlich
der Korrektur des entsprechenden Phänotyps selektionieren. Nach
Subclonierung und Sequenzierung des DNA-Fragments, das in der Lage
ist den Wildtyp-Phänotyp wieder
herzustellen, kann man dieses Fragment verwenden, um die Aktivität des Genprodukts
auszuschalten, das in P. pastoris von OCH1 codiert wird.
-
Falls
die gesamte genomische Sequenz eines bestimmten Pilzes von Interesse
bekannt ist, kann man alternativ solche Gene identifizieren, indem
man einfach öffentlich
erhältliche
DNA-Datenbanken durchsucht, die von einigen Quellen erhältlich sind,
wie NCBI, Swissprot etc. Indem man zum Beispiel eine vorgegebene genomische
Sequenz oder eine Datenbank mit einem bekannten 1,6-Mannosyltransferase-Gen
(OCH1) aus S. cerevisiae durchsucht, kann man in der Lage sein,
in einem solchen Genom Gene mit hoher Homologie zu identifizieren,
die mit einem hohen Grad an Sicherheit ein Gen codieren, das 1,6-Mannosyltransferase-Aktivität besitzt.
Unter Verwendung jedes dieser beiden Ansätze wurden Homologe zu einigen
bekannten Mannosyltransferasen von S. cerevisiae in P. pastoris
identifiziert. Diese Gene besitzen ähnliche Funktionen wie diejenigen
Gene, die bei der Mannosylierung von Proteinen in S. cerevisiae
beteiligt sind, und somit kann ihre Deletion dazu verwendet werden
das Glycosylierungs-Muster in P. pastoris oder einem beliebigen
anderen Pilz mit ähnlichen
Glycosylierungs-Signalwegen zu manipulieren.
-
Die
Erzeugung von Gen-knock outs, nachdem eine vorgegebene Ziel-Gensequenz bestimmt
worden ist, ist im Bereich der Hefe- und Pilzmolekularbiologie ein
gut etabliertes Verfahren und kann von jedem beliebigen durchschnittlichen
Fachmann durchgeführt
werden (R. Rothsteins, (1991) Methods in Enzymology, Bd, 194, S.
281). In der Tat kann die Wahl eines Wirts-Organismus durch die
Verfügbarkeit
von guten Transformations- und Gen-Zerstörungs-Verfahren für einen
solchen Wirt beeinflusst werden. Falls einige Mannosyltransferasen
einem Knock-out ausgesetzt werden müssen, erlaubt das Verfahren,
das von Alani und Kleckner entwickelt wurde, die wiederholte Verwendung
der URA3-Marker, um nacheinander sämtliche unerwünschte endogene
Mannosyltransferase-Aktivität
auszuschalten. Dieses Verfahren wurde durch andere verfeinert, beinhaltet
aber im Grundsatz die Verwendung von zwei sich wiederholenden DNA-Sequenzen,
die einen gegen selektierbaren Marker flankieren. Zum Beispiel:
URA3 kann als ein Marker verwendet werden, um die Selektion einer
Transformante zu gewährleisten,
die ein Konstrukt integriert hat. Durch Flankierung des URA3-Markers
mit direkten sich wiederholenden Sequenzen, kann man als erstes
nach Transformanten selektionieren, die das Konstrukt integriert
haben und somit das Zielgen zerstört haben. Nach Isolierung der
Transformanten und deren Charakterisierung kann man in einer zweiten
Runde nach denjenigen gegenselektionieren, die gegen 5' FOA resistent sind.
Kolonien, die in der Lage sind, auf Platten zu überleben, die 5' FOA enthalten, haben den
URA3-Marker durch ein Crossing over-Ereignis, an dem die vorstehend
erwähnten
sich wiederholenden Sequenzen beteiligt sind, wieder verloren. Dieser
Ansatz erlaubt somit die wiederholte Verwendung des gleichen Markers
und erleichtert die Zerstörung
von multiplen Genen, ohne zusätzliche
Marker zu benötigen.
-
Das
Ausschalten spezifischer Mannosyltransferasen wie 1,6-Mannosyltransferase
(OCH1), Mannosylphosphat-Transferasen (MNN4, MNN6 oder Gene, die
Ibd-Mutanten komplementieren) in P. pastoris erlaubt die Erzeugung
von gentechnisch veränderten
Stämmen
dieses Organismus, die vorrangig Man
8GlcNAc
2 synthetisieren und somit verwendet werden
können,
um das Glycosylierungs-Muster weiter zu modifizieren, um komplexeren
menschlichen Glycoform-Strukturen in stärkerem Maße ähnlich zu sein. Eine bevorzugte
Ausführungsform
dieses Verfahrens verwendet bekannte DNA-Sequenzen, die bekannte
biochemische Glycosylierungs-Aktivitäten codieren, um ähnliche
oder identische biochemische Funktionen in P. pastoris auszuschalten,
sodass die Glycosylierungs-Struktur des resultierenden gentechnisch
veränderten
P. pastoris-Stammes modifiziert ist. Tabelle
2.
Legende: M = A oder C, R = A oder G, W = A oder
T, S = C oder G, Y = C oder T, K = G oder T, V = A oder C oder G,
H = A oder C oder T, D = A oder G oder T, B = C oder G oder T, N
= G oder A oder T oder C.
-
Einfügung einer Mannosidase in den
gentechnisch veränderten
Wirt
-
Das
hierin beschriebene Verfahren ermöglicht es eine solche Struktur
mit hohem Ertrag zu erhalten, für
den Zweck, diese zu modifizieren, um komplexe N-Glycane zu erhalten. Ein erfolgreiches
Schema, um geeignete Man5GlcNAc2-Strukturen zu erhalten,
muss zwei parallel laufende Ansätze
beinhalten: (1) Das Verringern der endogenen Mannosyltransferase-Aktivität und (2)
das Entfernen von 1,2-α-Mannose
durch Mannosidasen, um hohe Spiegel an geeigneten Man5GlcNAc2-Strukturen
zu erhalten. Was dieses Verfahren von dem Stand der Technik unterscheidet
ist, dass es diese beiden Probleme direkt bewältigt. Wie die Arbeit von Chiba und
Mitarbeitern zeigt, kann man Man8GlcNAc2-Strukturen in S. cerevisiae in ein Man5GlcNAc2-Isomer reduzieren,
indem man das Vorliegen einer Pilz-Mannosidase von A. Saitoi durch gentechnische
Verfahren in das große
ER einbringt. Die Mängel
von deren Verfahren sind zweifach: (1) In der extrazellulären Glycoprotein-Fraktion
werden keine ausreichenden Mengen von Man5GlcNAc2 gebildet (10%), und (2) es ist nicht klar, dass
die in vivo gebildete Man5GlcNAc2-Struktur
in der Tat in der Lage ist, GlcNAc durch die Wirkung von GlcNAc-Transferase zu akzeptieren.
Falls einige Glycosylierungsstellen in einem gewünschten Protein vorliegen,
ergibt sich die Wahrscheinlichkeit (P), ein solches Protein in einer
korrekten Form zu erhalten, aus der Beziehung P = (F)n,
wobei n der Anzahl an Glycosylierungsstellen entspricht und F der
Fraktion an gewünschten
Glycoformen entspricht. Ein Glycoprotein mit drei Glycosylierungsstellen
würde eine
Wahrscheinlichkeit von 0,1% besitzen, die geeigneten Vorstufen für das Prozenieren
von komplexen und Hybrid-N-Glycanen an allen Glycosylierungsstellen
bereitzustellen, was den wirtschaftlichen Wert eines solchen Ansatzes
beschränkt.
-
Die
meisten Enzyme, die in dem ER und dem Golgi-Apparat von S. cerevisiae
aktiv sind, haben ein pH-Optimum, das zwischen 6,5 und 7,5 liegt
(siehe Tabelle 3). Alle vorherigen Ansätze, die Mannosylierung durch
die Wirkung von rekombinanten Mannosidasen zu reduzieren, haben
sich auf Enzyme konzentriert, die ein pH-Optimum um etwa pH 5,0 besitzen (Martinet
et al., 1998, und Chiba et al., 1998), obwohl die Aktivität dieser
Enzyme bei pH 7,0 sogar auf weniger als 10% reduziert ist und diese
somit höchstwahrscheinlich
an dem Ort ihrer Verwendung, dem ER und dem frühen Golgi von P. pastoris und
S. cerevisiae, nicht ausreichende Aktivität bereitstellen. Ein bevorzugtes
Verfahren verwendet eine α-Mannosidase
in vivo, wobei das pH-Optimum der Mannosidase nicht mehr als 1,4
pH-Einheiten vom durchschnittlichen pH-Optimum anderer repräsentativer
Marker-Enzyme abweicht, die in dem/den gleichen Organellen) lokalisiert
sind. Das pH-Optimum des Enzyms, das auf ein spezifischen Organell
abzielt, sollte mit dem pH-Optimum von anderen Enzymen übereinstimmen,
die in dem gleichen Organell gefunden werden, sodass die maximale
Aktivität
pro Einheit Enzym erhalten wird. Tabelle 3 fasst die Aktivität von Mannosidasen
aus verschiedenen Quellen und deren jeweilige pH-Optima zusammen.
Tabelle 4 fasst deren Lokalisation zusammen.
-
Tabelle
3. Mannosidasen und ihr pH-Optimum
-
Wenn
man versucht High-Mannose-Strukturen zu trimmen, um Man5GlcNAc2 in dem ER oder dem Golgi-Apparat von S.
cerevisiae zu erhalten, kann man ein beliebiges Enzym oder eine
Kombination von Enzymen auswählen,
das/die (1) ein ausreichend enges pH-Optimum hat haben (d.h. zwischen
pH 5,2 und pH 7,8), und (2) von dem/denen man weiß, dass
es/sie alleine oder gemeinsam die spezifische isomere Man5GlcNAc2-Struktur
erzeugt/erzeugen, die erforderlich ist, um das nachfolgende Hinzufügen von
GlcNAc durch GnTI zu gewährleisten.
Jedes beliebige Enzym oder jede beliebige Kombination von Enzymen,
das/die gezeigt hat/haben, dass es/sie eine Struktur erzeugt/erzeugen,
die in vitro durch GnTI in GlcNAcMan5GlcNAc2 umgewandelt werden kann, würde eine
geeignete Wahl darstellen. Diese Kenntnis kann aus der wissenschaftlichen
Literatur erhalten werden oder experimentell durch Bestimmen, dass
eine potenzielle Mannosidase Man8GlcNAc2-PA in Man5GlcNAc2-PA umwandeln kann, und dann Testen, ob
die erhaltene Man5GlcNAc2-PA-Struktur
als ein Substrat für
GnTI und UDP-GlcNAc dienen kann, um GlcNAcMan5GlcNAc2 in vitro zu erhalten. Mannosidase IA von
einer menschlichen oder murinen Quelle würde zum Beispiel eine geeignete
Wahl sein.
-
1,2-Mannosidase-Aktivität in dem
ER und dem Golgi
-
Vorherige
Versuche Mannosylierung durch die Wirkung von clonierten exogenen
Mannosidasen zu reduzieren sind daran gescheitert Glycoproteine
bereitzustellen, die eine ausreichende Fraktion (z.B. > 27 Mol%) an N-Glycanen
enthalten, die die Struktur Man5GlcNAc2 besitzen (Martinet et al., 1998, und Chiba
et al., 1998). Diese Enzyme sollten effizient in dem ER oder dem
Golgi-Apparat funktionieren, um effizient nascierende Glycoproteine
umzuwandeln. Während
die beiden Mannosidasen, die im Stand der Technik verwendet werden (aus
A. saitoi und T. reesei), ein pH-Optimum von 5,0 besitzen, haben
die meisten Enzyme, die in dem ER und dem Golgi-Apparat von Hefe
(z.B. S. cerevisiae) aktiv sind, pH-Optima, die zwischen 6,5 und
7,5 liegen (siehe Tabelle 3). Da die Glycosylierung von Proteinen
ein hoch entwickeltes und wirkungsvolles Verfahren darstellt, kann
geschlossen werden, dass der interne pH-Wert des ER und des Golgi
ebenfalls in dem Bereich von etwa 6–8 liegt. Bei pH 7,0 ist die
Aktivität
der Mannosidasen, die im Stand der Technik verwendet werden, auf
weniger als 10% reduziert, was für
die wirkungsvolle Man5GlcNAc2-Produktion
in vivo nicht ausreichend ist. Chibas Arbeit wurde auch in EP-A1
1 211 310 berichtet. Weitere Anmerkungen sind hierin vorstehend
bereitgestellt.
-
Tabelle
4. Zelluläre
Lokalisation und pH-Optima von verschiedenen mit Glycosylierung
in Zusammenhang stehenden Enzymen von S. cerevisiae.
-
Das α-1,2-Mannosidase-Enzym
sollte bei einem pH-Wert zwischen 5,1 und 8,0 optimale Aktivität besitzen.
In einer bevorzugten Ausführungsform
hat das Enzym eine optimale Aktivität bei einem pH-Wert zwischen
5,9 und 7,5. Der optimale pH-Wert
kann unter in vitro-Test-Bedingungen bestimmt werden. Bevorzugte Mannosidasen
beinhalten diejenigen, die in Tabelle 3 aufgelistet sind und die
geeignete pH-Optima besitzen, z.B. Aspergillus nidulans, Homo sapiens
IA (Golgi), Homo sapiens IB (Golgi), Lepidoptera-Insekten-Zellen (IPLB-SF21AE),
Homo sapiens, Maus-IB (Golgi) und Xanthomonas manihotis. In einer
bevorzugten Ausführungsform
wird ein einzelnes cloniertes Mannosidase-Gen in dem Wirtsorganismus
exprimiert. Es kann jedoch in einigen Fällen wünschenswert sein einige verschiedene
Mannosidase-Gene zu exprimieren oder einige Kopien eines bestimmten
Gens, um die angemessene Herstellung von Man5GlcNAc2 zu erreichen. In den Fällen, in denen multiple Gene
verwendet werden, sollten die codierten Mannosidasen alle pH-Optima
innerhalb des bevorzugten Bereiches von 5,1 bis 8,0 besitzen oder
insbesondere zwischen 5,9 und 7,5. In einer speziell bevorzugten
Ausführungsform
wird die Mannosidase-Aktivität
zu dem ER oder den cis-Golgi geleitet, wo die frühen Reaktionen der Glycosylierung
auftreten.
-
Bildung von komplexen
N-Glycanen
-
Ein
zweiter Schritt des Verfahrens beinhaltet das aufeinanderfolgende
Anhängen
von Zuckern an die nascierende Kohlenhydrat-Struktur durch Einbringen
der Expression von Glucosyltransferasen in den Golgi-Apparat. Dieser
Prozess benötigt
als erstes die funktionelle Expression von GnTI in dem frühen oder
medialen Golgi-Apparat sowie die Gewährleistung der Bereitstellung
von UDP-GlcNAc in ausreichenden Mengen.
-
Integrations-Stellen
-
Da
das ultimative Ziel dieses gentechnischen Manipulationsversuchs
ein robuster Stamm für
die Herstellung von Protein ist, der in der Lage ist dies in einem
industriellen Fermentations-Verfahren gut durchzuführen, beinhaltet
die Integration multipler Gene in das Pilz-Chromosom eine sorgfältige Planung.
Der veränderte
Stamm wird sehr wahrscheinlich mit einer Auswahl unterschiedlicher
Gene transformiert werden müssen und
diese Gene werden in einer stabilen Art und Weise transformiert
werden müssen,
um zu gewährleisten, dass
die gewünschte
Aktivität
während
des Fermentations-Prozesses erhalten bleibt. Jede beliebige Kombination
der nachfolgenden Enzym-Aktivitäten
wird in den Proteinexpressions-Pilzwirt eingebracht werden müssen: Sialyltransferasen,
Mannosidasen, Fucosyltransferasen, Galactosyltransferasen, Glucosyltransferasen, GlcNAc-Transferasen, ER-
und Golgi-spezifische Transporter (z.B. Syn- und Antiport-Transporter für UDP-Galactose
und andere Vorstufen), andere Enzyme, die an der Prozessierung von
Oligosacchariden beteiligt sind, und Enzyme, die bei der Synthese
von aktivierten Oligosaccharid-Vorstufen beteiligt sind, wie UDP-Galactose, CMP-N-Acetyl-Neuraminsäure. Gleichzeitig
wird eine Auswahl von Genen ausgeschaltet werden müssen, die Enzyme
codieren, von denen bekannt ist, dass sie für nicht-menschliche-Glycosylierungs-Reaktionen
charakteristisch sind.
-
Leiten von Glycosyltransferasen
zu spezifischen Organellen:
-
Glycosyltransferasen
und Mannosidasen kleiden die innere (luminale) Oberfläche des
ER und des Golgi-Apparates aus und stellen dabei eine "katalytische" Oberfläche bereit,
die die aufeinanderfolgende Prozessierung von Glycoproteinen erlaubt,
während
diese durch das ER und das Golgi-Netzwerk voranschreiten. In der
Tat stellen die multiplen Kompartimente des Cis-, des medialen und
des Trans-Golgi und das Trans-Golgi-Netzwerk (TGN) die veschiedenen
Orte bereit, in denen die geordnete Aufeinanderfolge von Glycosylierungs-Reaktionen
stattfinden kann. Während
ein Glycoprotein von der Synthese in dem ER zu seiner vollen Reife
in dem späten
Golgi oder dem TGN voranschreitet, ist es in aufeinanderfolgenden
Schritten veschiedenen Glycosidasen, Mannosidasen und Glycosyl-Transferasen
ausgesetzt, sodass eine spezifische Kohlenhydrat-Struktur synthetisiert
werden kann. Viel Arbeit wurde aufgewendet, um den genauen Mechanismus
zu offenbaren, durch den diese Enzyme in ihren jeweiligen Organellen
festgehalten und verankert werden. Das sich entwickelnde Bild ist
komplex, aber Hinweise legen nahe, dass die Stamm-Region, die Membrandurchdringende
Region und der cytoplasmatische Schwanz einzeln oder gemeinsam die
Enzyme an die Membran von einzelnen Organellen dirigieren und dabei
die assoziierte katalytische Domäne
an diesen Ort lokalisieren.
-
Zielsequenzen
sind gut bekannt und in der wissenschaftlichen Literatur und in öffentlichen
Datenbanken beschrieben, wie nachfolgend im Hinblick auf Bibliotheken
zur Auswahl von Zielsequenzen und zielgerichteten Enzymen ausführlicher
diskutiert.
-
Verfahren zur Herstellung
einer Bibliothek, um modifizierte Glycosylierungswege zu produzieren.
-
Eine
Bibliothek, die mindestens zwei Gene beinhaltet, die exogene Glycosylierungs-Enzyme
codieren, wird in den Wirtsorganismus transformiert, wobei eine
genetisch gemischte Population hergestellt wird. Transformanten,
die den gewünschten
Glycosylierungs-Phänotyp
besitzen, werden dann aus der gemischten Population selektioniert.
In einer bevorzugten Ausführungsform
ist der Wirtsorganismus eine Hefe, insbesondere P. pastoris, und
der Wirts-Glycosylierungsweg
wird durch die funktionelle Expression von einem oder mehreren menschlichen
oder tierischen Glycosylierungs-Enzymen modifiziert, wobei Protein-N-Glycane erhalten
werden, die menschlichen Glycoformen ähnlich oder mit diesen identisch
sind. In einer speziell bevorzugten Ausführungsform beinhaltet die DNA-Bibliothek genetische
Konstrukte, die Fusionen von Glycosylierungs-Enzymen mit Zielsequenzen
für verschiedene
zelluläre
Loci codieren, die an der Glycosylierung beteiligt sind, insbesondere
das ER, den Cis-Golgi, den medialen Golgi oder den Trans-Golgi.
-
Beispiele
von Modifikationen für
Glycosylierung, die unter Verwendung des Verfahrens durchgeführt werden
können,
sind: (1) die Veränderung
eines eukaryontischen Mikroorganismus derart, dass er Mannose-Reste
von Man8GlcNAc2 trimmt,
sodass Man5GlcNAc2 als
ein Protein-N-Glycan entsteht; (2) Veränderung eines eukaryontischen
Mikroorganismus derart, dass durch die Wirkung von GlcNAc- Transferase I ein
N-Acetyl-Glucosamin (GlcNAc)-Rest an Man5GlcNAc2 angehängt
wird; (3) Veränderung
eines eukaryontischen Mikroorganismus derart, dass er funktionell
ein Enzym wie eine N-Acetyl-Glucosamin-Transferase (GnTI, GnTII, GnTIII,
GnTIV, GnTV, GnTVI), eine Mannosidase II, eine Fucosyltransferase,
eine Galactosyltransferase (GalT) oder Sialyltransferasen (ST) exprimiert.
-
Durch
Wiederholen des Verfahrens können
zunehmend komplexe Glycosylierungswege in den Ziel-Mikroorganismus
eingebracht werden. In einer bevorzugten Ausführungsform wird der Wirtsorganismus zwei
oder mehrere Male mit DNA-Bibliotheken transformiert, die Sequenzen
enthalten, die Glycosylierungs-Aktivitäten codieren.
Nach jeder Runde der Transformation oder in einer alternativen Ausführungsform nachdem
einige Transformationen aufgetreten sind, kann die Selektionierung
der gewünschten
Phänotypen durchgeführt werden.
Auf diese Art und Weise können
komplexe Glycosylierungswege schnell eingefügt werden.
-
DNA-Bibliotheken
-
Es
ist erforderlich eine DNA-Bibliothek zusammenzustellen, die mindestens
zwei exogene Gene beinhaltet, die Glycosylierungs-Enzyme codieren.
Zusätzlich
zu den Sequenzen im offenen Leserahmen wird es im Allgemeinen bevorzugt
jedes Konstrukt der Bibliothek mit solchen Promotoren, Transkriptions-Terminatoren,
Enhancern, Ribosomen-Bindungsstellen und anderen funktionellen Sequenzen
bereitzustellen, wie es notwendig ist, um nach Transformation in
den Wirtsorganismus eine effektive Transkription und Translation
der Gene zu gewährleisten.
Dort, wo der Wirt Pichia pastoris ist, beinhalten geeignete Promotoren
zum Beispiel die AOX1-, AOX2-, DAS- und P40-Promotoren. Es wird
auch bevorzugt, jedes Konstrukt mit mindestens einem selektionierbaren
Marker bereitzustellen, wie ein Gen, das eine Arzneistoff-Resistenz
verleiht oder das einen Defekt im Wirts-Metabolismus komplementiert.
Das Vorliegen des Markers ist für
die nachfolgende Selektion von Transformanten zweckmäßig; in
Hefe können
zum Beispiel die URA3-, HIS4-, SUC2-, G418-, BLA- oder SH BLE-Gene
verwendet werden.
-
In
einigen Fällen
kann die Bibliothek direkt aus existierenden oder Wildtyp-Genen zusammengesetzt werden.
In einer bevorzugten Ausführungsform
wird die DNA-Bibliothek jedoch aus der Fusion von zwei oder mehreren
Unter-Bibliotheken zusammengesetzt. Durch die Ligierung der Unter-Bibliotheken
im Leserahmen ist es möglich
eine große
Anzahl von neuen genetischen Konstrukten zu erzeugen, die zweckmäßige zielgerichtete
Glycosylierungs-Aktivitäten
codieren. Eine zweckmäßige Unter-Bibliothek
beinhaltet zum Beispiel DNA-Sequenzen, die irgendeine beliebige
Kombination von Enzymen wie Sialyltransferasen, Mannosidasen, Fucosyltransferasen,
Galactosyltransferasen, Glucosyltransferasen und GlcNAc-Transferasen codieren.
Vorzugsweise sind die Enzyme von menschlichem Ursprung, obwohl andere
Säuger-,
tierische oder Pilz-Enzyme ebenfalls zweckmäßig sind. In einer bevorzugten
Ausführungsform
sind die Gene verkürzt,
wobei Fragmente erhalten werden, die die katalytischen Domänen der
Enzyme codieren. Durch das Entfernen der endogenen Ziel-Sequenzen
können
die Enzyme dann von neuem ausgerichtet und an anderen zellulären Orten
exprimiert werden. Die Auswahl solcher katalytischer Domänen kann
durch die Kenntnis der jeweiligen Umgebung geleitet werden, in der
die katalytische Domäne
im Nachfolgenden aktiv sein soll. Wenn ein bestimmtes Glycosylierungs-Enzym
zum Beispiel in dem späten
Golgi aktiv sein soll und alle bekannten Enzyme des Wirtsorganismus
in dem späten
Golgi ein bestimmtes pH-Optimum besitzen, dann wird eine katalytische
Domäne
ausgewählt,
die bei diesem pH-Wert eine angemessene Aktivität besitzt.
-
Eine
andere zweckmäßige Unter-Bibliothek
beinhaltet DNA-Sequenzen, die Signalpeptide codieren, die in der
Lokalisation eines Proteins an einen bestimmten Ort innerhalb des
ER, des Golgi oder des Trans-Golgi-Netzwerks resultieren. Diese
Signalsequenzen können
aus dem Wirtsorganismus sowie von anderen verwandten oder nicht
verwandten Organismen ausgewählt
werden. Membran-gebundene Proteine des ER oder des Golgi können typischerweise
zum Beispiel N-terminale Sequenzen beinhalten, die einen cytosolischen
Schwanz (ct), eine Transmembran-Domäne (tmd)
und eine Stamm-Region (sr) codieren. Die ct-, tmd- und sr-Sequenzen sind ausreichend,
einzeln oder in Kombination, um Proteine an die innere (luminale)
Membran des Organells zu verankern. Entsprechend beinhaltet eine
bevorzugte Ausführungsform
der Unter-Bibliothek von Signalsequenzen ct-, tmd- und/oder sr-Sequenzen
dieser Proteine. In einigen Fällen
ist es wünschenswert
die Unter-Bibliothek mit unterschiedlichen Längen der sr-Sequenz bereitzustellen.
Dies kann durch PCR erreicht werden, wobei Primer verwendet werden,
die an das 5'-Ende
der DNA binden, die die cytosolische Region codiert, und wobei eine
Serie von gegenläufigen
Primern verwendet wird, die an verschiedene Stellen der Stammregion
binden. Noch andere zweckmäßige Quellen
von Signalsequenzen beinhalten Wiederfindungs-Signalpeptide, z.B.
die Tetrapeptide HDEL oder KDEL, die im Allgemeinen an dem C-terminalen Ende
von Proteinen gefunden werden, die retrograd in das ER oder den
Golgi transportiert werden. Noch andere Quellen von Signalsequenzen
beinhalten (a) Typ II-Membranproteine, (b) die Enzyme, die in Tabelle
3 aufgelistet sind, (c) Membran-durchdringende Nucleotid-Zucker-Transporter, die
in dem Golgi lokalisiert sind, und (d) Sequenzen, auf die sich in
Tabelle 5 bezogen wird.
-
Tabelle
5. Quellen zweckmäßiger Zielsequenzen
für Kompartimente
-
In
jedem Fall wird es stark bevorzugt, dass Signalsequenzen ausgewählt werden,
die für
die enzymatische(n) Aktivität
oder Aktivitäten
geeignet sind, die in dem Wirt eingefügt werden sollen. Für die Entwicklung eines
modifizierten Mikroorganismus, der in der Lage ist nascente N-Glycane
terminal zu sialysieren, ein Prozess, der in Menschen in dem späten Golgi
auftritt, ist es zum Beispiel wünschenswert
eine Unter-Bibliothek von Signalsequenzen zu verwenden, die von
späten
Golgi-Proteinen abstammt. In ähnlicher
Weise ist das Trimmen von Man8GlcNAc2 durch eine α-1,2-Mannosidase zum Erhalt
von Man5GlcNAc2 in
Menschen ein früher
Schritt bei der komplexen N-Glycan-Bildung. Es ist deshalb wünschenswert,
dass man erreicht, dass diese Umsetzung in dem ER oder frühen Golgi
eines manipulierten Wirtsorganismus auftritt. Es wird eine Unter-Bibliothek
verwendet, die ER- und frühe
Golgi-Zurückhaltesignale
codiert.
-
In
einer bevorzugten Ausführungsform
wird dann durch die Ligierung einer Unter-Bibliothek, die DNA enthält, die
Signalsequenzen codiert, mit einer Unter-Bibliothek, die DNA enthält, die
Glycosylierungs-Enzyme oder katalytisch aktive Fragmente davon codiert,
im Leserahmen eine DNA-Bibliothek konstruiert. Die resultierende
Bibliothek beinhaltet synthetische Gene, die Fusionsproteine codieren.
In einigen Fällen
ist es wünschenswert
an dem N-terminalen Ende eines Fusionsproteins oder in anderen Fällen am
C-terminalen Ende eine Signalsequenz bereitzustellen. In einigen
Fällen
können
Signalsequenzen innerhalb des offenen Leserahmens eines Enzyms eingefügt werden,
vorausgesetzt die Proteinstruktur individuell gefalteter Domänen wird
nicht zerstört.
-
Das
Verfahren ist am wirkungsvollsten wenn eine DNA-Bibliothek, die
in den Wirt transformiert wird, eine große Vielfalt von Sequenzen enthält, wobei
die Wahrscheinlichkeit erhöht
wird, dass mindestens eine Transformante den gewünschten Phänotyp zeigen wird. Entsprechend
kann eine DNA-Bibliothek oder eine bestehende Unter-Bibliothek vor
der Transformation einer oder mehreren Runden eines Gen-Shuffling,
einer fehlergeneigten PCR oder in vitro-Mutagenese unterzogen werden.
-
Transformation
-
Die
DNA-Bibliothek wird dann in den Wirtsorganismus transformiert. In
Hefe kann jedes beliebige geeignete Verfahren des DNA-Transfers
verwendet werden, wie Elektroporation, das Lithiumchlorid-Verfahren oder
das Sphäroplasten-Verfahren.
Um einen stabilen Stamm zu produzieren, der zur Fermentation in
hoher Dichte geeignet ist, ist es wünschenswert die DNA-Konstrukte
der Bibliothek in das Wirtsgenom zu integrieren. In einer bevorzugten
Ausführungsform
erfolgt die Integration über
homologe Rekombination, wobei Verfahren verwendet werden, die im
Stand der Technik bekannt sind. Es werden zum Beispiel DNA-Elemente
der Bibliothek mit flankierenden Sequenzen bereitgestellt, die homolog
sind zu Sequenzen des Wirtsorganismus. Auf diese Art und Weise findet
die Integration an einer bestimmten Stelle in dem Wirtsgenom statt,
ohne erwünschte
oder notwendige Gene zu zerstören.
In einer speziell bevorzugten Ausführungsform wird die DNA der
Bibliothek an die Stelle eines unerwünschten Gens in einem Wirtschromosom integriert,
was das Zerstören
oder die Deletion des Gens bewirkt. Zum Beispiel erlaubt die Integration
an die Stellen der OCH1-, MNN1- oder MNN4-Gene die Expression der
gewünschten
DNA der Bibliothek, während
die Expression von Enzymen verhindert wird, die bei der Hefe-Hypermannosylierung
von Glycoproteinen beteiligt sind. In anderen Ausführungsformen
kann die DNA der Bibliothek mit Hilfe eines Chromosoms, Plasmids,
retroviralen Vektors oder durch zufällige Integration in das Wirtsgenom
in den Wirt eingebracht werden. In jedem Fall ist es im Allgemeinen
wünschenswert
mit jedem DNA-Konstrukt der Bibliothek mindestens ein selektionierbares
Markergen einzuschließen,
das die leichte Selektion von Wirtsorganismen erlaubt, die stabil
transformiert worden sind. Wiederverwendbare Marker-Gene wie ura3,
auf oder gegen das selektioniert werden kann, sind speziell geeignet.
-
Selektionsverfahren
-
Nach
der Transformation des Wirtsstammes mit der DNA-Bibliothek werden
Transformanten ausgewählt,
die den gewünschten
Glycosylierungs-Phänotyp
zeigen. Die Selektion kann in einem einzelnen Schritt oder durch
eine Reihe phänotypischer
Anreicherungs- und/oder Verarmungs-Schritte durchgeführt werden, wobei
jeder beliebige aus einer Vielzahl von Tests oder Nachweisverfahren
verwendet werden kann. Die phänotypische
Charakterisierung kann von Hand durchgeführt werden oder einen automatischen
Hochdurchsatz-Durchmusterungs-Apparat
verwenden. Im Allgemeinen zeigt ein Wirtsorganismus Protein-N-Glycane
auf der Zelloberfläche,
wo verschiedene Glycoproteine lokalisiert sind. Entsprechend können intakte
Zellen nach einem gewünschten
Glycosylierungs-Phänotyp
durchmustert werden, indem die Zellen einem Lektin oder Antikörper ausgesetzt
werden, der spezifisch an das gewünschte N-Glycan bindet. Eine
große
Bandbreite Oligosaccharid-spezifischer Lektine ist kommerziell erhältlich (EY
Laboratories, San Mateo, CA). Alternativ dazu sind Antikörper gegen
spezifische menschliche oder tierische N-Glycane kommerziell erhältlich oder
können unter
Verwendung von Standard-Verfahren hergestellt werden. Ein geeignetes
Lektin oder ein geeignerter Antikörper kann mit einem Reporter-Molekül wie einem
Chromophor, Fluorophor, einem Radioisotop oder einem Enzym, das
ein chromogenes Substrat hat, konjugiert sein (Guillen et al., 1998.
Proc. Natl. Acad. Sci. USA 95 (14): 7888–7892). Die Durchmusterung
kann dann durchgeführt
werden, indem analytische Verfahren wie Spektrophotometrie, Fluorimetrie,
Fluoresenz-aktivierte Zell-Sortierung oder Scintillationszählung verwendet werden.
In anderen Fällen
kann es notwendig sein isolierte Glycoproteine oder N-Glycane von
transformierten Zellen zu analysieren. Die Protein-Isolierung kann
unter Verwendung von im Stand der Technik bekannten Verfahren durchgeführt werden.
In den Fällen,
in denen ein isoliertes N-Glycan erforderlich ist, kann ein Enzym wie
eine Endo-β-N-Acetyl-Glucosaminidase
(Genzyme Co., Boston, MA) verwendet werden, um die N-Glycane von
Glycoproteinen zu spalten. Isolierte Proteine oder N-Glycane können dann
durch Flüssigkeits-Chromatographie
(z.B. HPLC), Massen-Spektroskopie oder andere geeignete Verfahren
analysiert werden. Das US-Patent Nr. 5,595,900 lehrt einige Verfahren,
mit denen Zellen mit gewünschten
extrazellulären
Kohlenhydrat-Strukturen
identifiziert werden können.
Vor der Auswahl einer gewünschten
Transformante kann es wünschenswert
sein, die transformierte Population um diejenigen Zellen zu verarmen,
die unerwünschte
Phänotypen
besitzen. Wenn das Verfahren zum Beispiel verwendet wird, um eine
funktionelle Mannosidase-Aktivität in
Zellen einzubringen, wird die gewünschte Transformante niedrigere
Spiegel an Mannose im zellulärem
Glycoprotein besitzen. Exponiert man die transformierte Population
mit einem letalen Radioisotop von Mannose im Medium, verarmt man
die Population um Transformanten, die den ungewünschten Phänotyp besitzen, d.h. hohe Spiegel
an eingebauter Mannose. Alternativ dazu kann ein cytotoxisches Lektin
oder ein cytotoxischer Antikörper
verwendet werden, das/der gegen ein unerwünschtes N-Glycan gerichtet
ist, um eine transformierte Population um unerwünschte Phänotypen zu verarmen.
-
Verfahren zur Bereitstellung
von Zucker-Nucleotid-Vorläufern
an den Golgi-Apparat
-
Für eine Glycosyltransferase,
die in dem Golgi zufriedenstellend funktionieren soll, ist es notwendig, dass
dem Enzym eine ausreichende Konzentration eines geeigneten Nucleotid-Zuckers
bereitgestellt wird, der den Hoch-Energie-Spender der Zuckereinheit
darstellt, die an ein nascierendes Glycoprotein angefügt wird. Diese
Nucleotid-Zucker werden dem geeigneten Kompartiment bereitgestellt,
indem ein exogenes Gen, das einen Zucker-Nucleotid-Transporter codiert,
in dem Wirts-Mikroorganismus
exprimiert wird. Die Wahl des Transporter-Enzyms wird durch die
Natur der verwendeten exogenen Glycosyltransferase beeinflusst.
Eine GlcNAc-Transferase
zum Beispiel kann einen UDP-GlcNAc-Transporter benötigen, eine Fucosyltransferase kann
einen GDP-Fucose-Transporter benötigen,
eine Galactosyltransferase kann einen UDP-Galactose-Transporter
benötigen
oder eine Sialyltransferase kann einen CMP-Sialinsäure-Transporter
benötigen.
-
Das
zugegebene Transporter-Protein befördert einen Nucleotid-Zucker
vom Cytosol in den Golgi-Apparat, wo der Nucleotid-Zucker von der
Glycosyltransferase umgesetzt wird, z.B. um ein N-Glycan zu verlängern. Die
Umsetzung setzt ein Nucleosid-Diphosphat oder -Monophosphat frei,
z.B. UDP, GDP oder CMP. Da die Ansammlung eines Nucleosid-Diphosphats
die weitere Aktivität
einer Glycosyltransferase hemmt, ist es häufig auch wünschenswert eine exprimierte
Kopie eines Gens bereitzustellen, das eine Nucleotid-Diphosphatase
codiert. Die Diphosphatase (spezifisch für UDP oder GDP, soweit angemessen)
hydrolysiert das Diphosphonucleosid, wobei ein Nucleosid-Monophosphat
und anorganisches Phosphat erhalten werden. Das Nucleosid-Monophosphat
hemmt die Glycotransferase nicht und wird in jedem Fall durch ein
endogenes zelluläres System
aus dem Golgi exportiert. Geeignete Transporter-Enzyme, die typischerweise
aus einem Säuger
stammen, sind nachfolgend beschrieben.
-
Beispiele
-
Die
Verwendung des vorstehenden allgemeinen Verfahrens kann durch Bezugnahme
auf die nachfolgenden nicht einschränkenden Beispiele verstanden
werden. Beispiele von bevorzugten Ausführungsformen sind auch in Tabelle
6 zusammengefasst.
-
Beispiel 1: Manipulation
von P. pastoris mit α-1,2-Mannosidase,
um Interferon herzustellen.
-
Eine α-1,2-Mannosidase
ist erforderlich, um Man8GlcNAc2 zu
trimmen, wobei Man5GlcNAc2 erhalten wird,
ein notwendiges Zwischenprodukt für die Bildung eines komplexen
N-Glycans. Eine OCH1-Mutante von P. pastoris wird manipuliert, um
menschliches Interferon-β unter
der Kontrolle eines aox-Promotors zu exprimieren. Durch Ligierung
der katalytischen Domäne
von menschlicher Mannosidase IB (eine α-1,2-Mannosidase) im Leserahmen
mit einer Unter-Bibliothek, die Sequenzen beinhaltet, die frühe Golgi-Lokalisierungs-Peptide
codieren, wird eine DNA-Bibliothek konstruiert. Die DNA-Bibliothek
wird dann in den Wirtsorganismus transformiert, wobei eine genetisch
gemischte Population resultiert, in der einzelne Transformanten
sowohl Interferon-β als
auch ein synthetisches Mannosidase-Gen aus der Bibliothek exprimieren.
Einzelne Transformanten-Kolonien werden gezüchtet und die Herstellung von
Interferon wird durch die Zugabe von Methanol induziert. Unter diesen
Bedingungen beinhalten mehr als 90% des sekretierten Proteins Interferon-β. Die Überstande
werden durch C18-Silica-Umkehrphasen-Chromatographie gereinigt,
um Salze und niedermolekulare Fremdstoffe zu entfernen. Gewünschte Transformanten,
die angemessen zielgerichtete aktive α-1,2-Mannosidase exprimieren,
stellen Interferon-β einschließlich N-Glycanen
der Struktur Man5GlcNAc2 her,
die verglichen mit dem Interferon des parentalen Stammes eine reduzierte
molekulare Masse besitzen. Die gereinigten Überstande, einschließlich Interferon-β, werden
durch MALDI-TOF- Massenspektroskopie analysiert und Kolonien werden
identifiziert, die die gewünschte
Form von Interferon-β exprimieren.
-
Beispiel 2: Manipulation
eines Stammes zur Expression von GlcNAc-Transferase I
-
GlcNAc-Transferase
I-Aktivität
ist für
die Reifung von komplexen N-Glycanen erforderlich. Man5GlcNAc2 kann ausschließlich durch Mannosidase II
getrimmt werden, ein notwendiger Schritt bei der Bildung menschlicher
Glycoformen nach dem Anhängen
von GlcNAc an den terminalen α-1,3-Mannose-Rest durch
GlcNAc-Transferase
I (Schachter, 1991 Glycobiology 1(5): 453–461). Entsprechend wird eine
Bibliothek hergestellt, die DNA-Fragmente enthält, die geeignete zielgerichtete
GlcNAc-Transferase 1-Gene codieren. Der Wirtsorganismus ist ein
Stamm, z.B. eine Hefe, der für
Hypermannosylierung defizient ist (z.B. eine OCH1-Mutante), das
Substrat UDP-GlcNAc in dem Golgi und/oder dem ER bereitstellt und
N-Glycane der Struktur Man5GlcNAc2 in dem Golgi und/oder dem ER bereitstellt.
Nach Transformation des Wirts mit der DNA-Bibliothek werden die
Transformanten nach denjenigen durchmustert, die die höchste Konzentration
an terminalem GlcNAc auf der Zell-Oberfläche besitzen oder alternativ
dazu das Protein sekretieren, das den höchsten terminalen GLcNAc-Gehalt
besitzt. Eine solche Durchmusterung wird unter Verwendung eines
visuellen Verfahrens (z.B. eine Färbemethode), eines spezifischen
terminalen GLcNAc-bindenden Antikörpers oder eines Lektins durchgeführt. Alternativ
dazu zeigen die gewünschten
Transformanten eine reduzierte Bindung bestimmter Lektine, die für terminale
Mannose-Reste spezifisch sind.
-
Beispiel 3: Manipulation
von Stämmen
mit einer Mannosidase II
-
In
einem anderen Beispiel ist es erwünscht die zwei verbleibenden
terminalen Mannosen aus der Struktur GlcNAcMan5GlcNAc2 durch Wirkung einer Mannosidase II zu entfernen,
um eine menschliche Glycoform in einem Mikroorganismus zu erzeugen.
Eine DNA-Bibliothek, die Sequenzen beinhaltet, die Cis- und mediale
Golgi-Lokalisations-Signale codieren, wird im Leserahmen mit einer
Bibliothek fusioniert, die katalytische Domänen von Mannosidase II codiert.
Der Wirtsorganismus ist ein Stamm, z.B. eine Hefe, der für Hypermannosylierung
defizient ist (z.B. eine OCH1-Mutante) und N-Glycane bereitstellt,
die die Struktur GlcNAcMan5GlcNAc2 in dem Golgi und/oder dem ER besitzen.
Nach Transformation werden Organismen selektioniert, die den gewünschten
Glycosylierungs-Phänotyp
besitzen. In einem Verfahren wird ein in vitro-Test verwendet. Die
gewünschte
Struktur GlcNAcMan3GlcNAc2 (jedoch
nicht unerwünschtes
GlcNAcMan5GlcNAc2) ist
ein Substrat für
das Enzym GlcNAc-Transferase II. Entsprechend können einzelne Kolonien unter
Verwendung dieses Enzyms in vitro in Anwesenheit des Substrats,
UDP-GlcNAc, getestet werden. Die Freisetzung von UDP wird entweder
durch HPLC oder einen enzymatischen Test für UDP bestimmt. In einer alternativen Ausführungsform
wird radioaktiv markiertes UDP-GlcNAc verwendet.
-
Die
vorangehenden in vitro-Tests werden in geeigneter Weise mit einzelnen
Kolonien durchgeführt, wobei
eine Hochdurchsatz-Durchmusterungs-Apparatur verwendet wird. In
einer alternativen Ausführungsform
wird ein Lektin-Bindungstest verwendet. In diesem Fall erlaubt die
reduzierte Bindung von Lektinen, die für terminale Mannosen spezifisch
sind, die Selektion von Transformanten, die den gewünschten
Phänotyp
besitzen. Galantus nivalis-Lektin zum Beispiel bindet spezifisch
an terminale α-1,3-Mannose,
deren Konzentration in Anwesenheit von funktionell exprimierter
Mannosidase II-Aktivität
reduziert ist. In einem geeigneten Verfahren wird G. nivalis-Lektin
verwendet, das an einen festen Agarose-Träger (erhältlich von Sigma Chemical, St.
Louis, MO) angeheftet ist, um die transformierte Population an Zellen
zu verarmen, die hohe Spiegel an terminaler α-1,3-Mannose aufweisen.
-
Beispiel 4: Manipulation
von Organismen, die Sialyltransferase exprimieren
-
Die
Enzyme α-2,3-Sialyltransferase
und α-2,6-Sialyltransferase
fügen terminale
Sialinsäure
an Galactose-Reste in nascierenden menschlichen N-Glycanen an, was
zu reifen Glycoproteinen führt.
Im Menschen treten die Reaktionen in dem Trans-Golgi oder dem TGN
auf. Entsprechend wird durch die Fusion von Sequenzen, die katalytische
Domänen
von Sialyltransferase codieren, im Leserahmen mit Sequenzen, die
Trans-Golgi- oder TGN-Lokalisierungs-Signale codieren, eine DNA-Bibliothek
erzeugt. Der Wirtsorganismus ist ein Stamm, z.B. eine Hefe, der
für Hypermannosylierung
defizient ist (z.B. OCH1-Mutante), die N-Glycane bereitstellt, die terminate
Galactose-Reste in dem Trans-Golgi oder dem TGN besitzen, und eine
ausreichende Konzentration an CMP-Sialinsäure in dem Trans-Golgi oder
TGN bereitstellt. Nach der Transformation werden Transformanten
selektioniert, die den gewünschten
Phänotyp
besitzen, wobei ein fluoreszierender Antikörper verwendet wird, der für N-Glycane
spezifisch ist, die eine terminale Sialinsäure besitzen.
-
Beispiel 5: Verfahren
zur Manipulation von Stämmen,
um einen UDP-GlcNAc-Transporter
zu exprimieren
-
Die
cDNA von menschlichem Golgi-UDP-GlcNAc-Transporter wurde von Ishida
und Mitarbeitern cloniert (Ishida; N. et al. 1999 J. Biochem. 126
(1): 68–77).
Guillen und Mitarbeiter haben den Hunde-Nieren-Golgi-UDP-GlcNAc-Transporter
durch phänotypische
Korrektur einer Kluyveromyces lactis-Mutante cloniert, die für den Golgi-UDP-GlcNAc-Transport
defizient war (Guillen, E. et al. 1998). So besitzt ein Säuger-Golgi-UDP-GlcNAc-Transporter-Gen
alle notwendigen Informationen dass das Protein funktionell exprimiert
und zu den Golgi-Apparat von Hefe gelenkt wird.
-
Beispiel 6: Verfahren
zur Manipulation von Stämmen,
um einen GDP-Fucose-Transporter
zu exprimieren
-
Der
Rattenleber-Golgi-Membran-GDP-Fucose-Transporter wurde von Puglielle,
L. und C. B. Hirschberg 1999 J. Biol. Chem. 274 (59): 35596–35600 identifiziert
und aufgereinigt. Das entsprechende Gen kann unter Verwendung von
Standard-Verfahren wie N-terminale Sequenzierung und Southern-Blottverfahren unter Verwendung
einer degenerierten DNA-Probe identifiziert werden. Das intakte
Gen wird dann in einem Wirtsorganismus exprimiert, der ebenfalls
eine Fucosyltransferase exprimiert.
-
Beispiel 7: Verfahren
zur Manipulation von Stämmen,
um einen UDP-Galactose-Transporter
zu exprimieren
-
Ein
menschlicher UDP-Galactose (UDP-Gal)-Transporter wurde cloniert,
und es wurde gezeigt, dass dieser in S. cerevisiae aktiv ist (Kainuma,
M., et al. 1999 Glycobiology 9 (2): 133–141). Ein zweiter menschlicher
UDP-Galactose-Transporter (hUGT1) wurde cloniert und in Ovarzellen
des Chinesischen Hamsters funktionell exprimiert. Aoki, K., et al.
1999 J. Biochem. 126 (5): 940–950.
Auf ähnliche
Weise haben Segawa und Mitarbeiter einen UDP-Galactose-Transporter
von Schizosaccharomyces pombe cloniert (Segawa, H., et al. 1999
Febs Letters 451 (3): 295–298).
-
CMP-Sialinsäure-Transporter
-
Ein
menschlicher CMP-Sialinsäure-Transporter
(hCST) wurde von Aoki und Mitarbeitern (1999) cloniert und in Lec
8 CHO-Zellen exprimiert. Die molekulare Clonierung des Hamster-CMP-Sialinsäure-Transporters
wurde ebenfalls erreicht (Eckhardt and Gerardy Schahn 1997 Eur.
J. Biochem. 248 (1): 187–192).
Die funktionelle Expression des murinen CMP-Sialinsäure-Transporters
wurde von Berninsone, P., et al. 1997 J. Biol. Chem. 272 (19): 12616–12619 in
Saccharomyces cerevisiae erreicht.
-
Tabelle
6. Beispiele für
bevorzugte Ausführungsformen
des Verfahrens zur Modifikation von Glycosylierung in einem eukaryontischen
Mikroorganismus, z.B. Pichia pastoris
-
-
Tabelle 7: Quellen für DNA- und
Protein-Sequenzen
-
- 1. European Bioinformatics Institute (EBI) ist ein Zentrum
für Forschung
und Dienstleistungen in der Bioinformatik: http://www.ebi.ac.uk/
- 2. Swissport Datenbank: http://www.expasy.ch/spr
- 3. Liste bekannter Glycosyltransferasen und deren Ursprung.
-
β1,2-(GnT I) EC 2.4.1.101
-
- 4. Menschliche cDNA, Kumar et al. (1990) Proc. Natl. Acad.
Sci. USA 87: 9948–9952
- 5. Menschliches Gen, Hull et al. (1991) Biochem. Biophys. Res.
Commun. 176: 608–615
- 6. Maus-cDNA, Kumar et al. (1992) Glycobiology 2: 383–393
- 7. Maus-Gen, Pownall et al. (1992) Genomics 12: 699–704
- 8. Maus-Gen (5'-flankierend,
nicht codierend), Yang et al. (1994) Glycobiology 5: 703–712
- 9. Kaninchen-cDNA, Sarkar et al. (1991) Proc. Natl. Acad. Sci.
USA 88: 234–238
- 10. Ratten-cDNA, Fukuda et al. (1994) Biosci. Biotechnol. Biochem.
58: 200–201
-
1,2-(GnT II) EC 2.4.1.143
-
- 11. Menschliches Gen, Tan et al. (1995) Eur. J. Biochem.
231: 317–328
- 12. Ratten-cDNA, D'Agostaro
et al. (1995) J. Biol. Chem. 270: 15211–15221
- 13. β1,4
(GnT III), EC 2.4.1.144
- 14. Menschliche cDNA, Ihara et al. (1993) J. Biochem. 113: 692–698
- 15. murines Gen, Bhaumik et al. (1995) Gene 164: 295–300
- 16. Ratten-cDNA, Nishikawa et al. (1992) J. Biol. Chem. 267:
18199–18204
-
β1,4-(GnT IV) EC 2.4.1.145
-
- 17. Menschliche cDNA, Yoshida et al. (1998) Glycoconjugate
Journal 15: 1115–1123
- 18. Rinder-cDNA, Minowa et al., Europäisches Patent EP 0 905 232 (31,6 (GnT V) EC 2.4.1.155
- 19. Menschliche cDNA, Saito et al. (1994) Biochem. Biophys.
Res. Commun. 198: 318–327
- 20. Ratten-cDNA, Shoreibah et al. (1993) J. Biol. Chem. 268:
15381–15385
-
β1,4-Galactosyltransferase, EC
2.4.1.90 (LacNAc-Synthetase) EC
-
2.4.1.22 (Lactose-Synthetase)
-
- 21. Rinder-cDNA, D'Agostaro
et al. (1989) Eur. J. Biochem. 183: 211–217
- 22. Rinder-cDNA (partiell), Narimatsu et al. (1986) Proc. Natl.
Acad. Sci. USA 83: 4720–4724
- 23. Rinder-cDNA (partiell), Masibay & Qasba (1989) Proc. Natl. Acad. Sci.
USA 86: 5733–5377
- 24. Rinder-cDNA (5'-Ende),
Russo et al. (1990) J. Biol. Chem. 265: 3324
- 25. Huhn-cDNA (partiell), Ghosh et al. (1992) Biochem. Biophys.
Res. Commun. 1215–1222
- 26. Menschliche cDNA, Masri et al. (1988) Biochem. Biophys.
Res. Commun. 157: 657–663
- 27. Menschliche cDNA, (HeLa-Zellen). Watzele & Berger (1990)
Nucl. Acids Res. 18: 7174
- 28. Menschliche cDNA, (partiell) Uejima et al (1992) Cancer
Res. 52: 6158–6163
- 29. Menschliche cDNA, (Karzinom) Appert et al. (1986) Biochem.
Biophys. Res. Commun. 139: 163–168
- 30. Menschliches Gen, Mengle-Gaw et al. (1991) Biochem. Biophys.
Res. Commun. 176: 1269–1276
- 31. murine cDNA, Nakazawa et al. (1988) J. Biochem. 104: 165–168
- 32. murine cDNA, Shaper et al. (1988) J. Biol. Chem. 263: 10420–10428
- 33. murine cDNA (neu), Uehara & Muramatsu, nicht veröffentlicht
- 34. murines Gen, Hollis et al. (1989) Biochem. Biophys. Res.
Commun. 162: 1069–1075
- 35. Rattenprotein (partiell), Bendiak et al. (1993) Eur. J.
Biochem. 216: 405–417
-
2,3-Sialyltransferase,
(ST3Gal II) (N-gekoppelt) (Gal-1,3/4-GlcNAc) EC 2.4.99.6
-
- 36. Menschliche cDNA, Kitagawa & Paulson (1993) Biochem. Biophys.
Res. Commun. 194: 375–382
- 37. Ratten-cDNA, Wen et al. (1992) J. Biol. Chem. 267: 21011–21019
-
2,6-Sialyltransferase,
(ST6Gal I) EC 2.4.99.1
-
- 38. Huhn, Kurosawa et al. (1994) Eur. J. Biochem 219: 375–381
- 39. Menschliche cDNA (partiell), Lance et al. (1989) Biochem.
Biophys. Res. Commun. 164: 225–232
- 40. Menschliche cDNA, Grundmann et al. (1990) Nucl. Acids Res.
18: 667
- 41. Menschliche cDNA, Zettlmeisl et al. (1992) Patent EPO475354-A/3
- 42. Menschliche cDNA, Stamenkovic et al. (1990) J. Exp. Med.
172: 641–643
(CD75)
- 43. Menschliche cDNA, Bast et al. (1992) J. Cell Biol. 116:
423–435
- 44. Menschliches Gen (partiell), Wang et al. (1993) J. Biol.
Chem. 268: 4355–4361
- 45. Menschliches Gen (5'-flankierend),
Aasheim et al. (1993) Eur. J. Biochem. 213: 467–475
- 46. Menschliches Gen (Promotor), Aas-Eng et al. (1995) Biochim.
Biophys. Acta 1261: 166–169
- 47. Maus-cDNA, Hamamoto et al. (1993) Bioorg. Med. Chem. 1:
141–145
- 48. Ratten-cDNA, Weinstein et al. (1987) J. Biol. Chem. 262:
17735–17743
- 49. Ratten-cDNA (Transkript-Fragmente), Wang et al. (1991) Glycobiology
1: 25–31,
Wang et al. (1990) J. Biol. Chem. 265: 17849–17853
- 50. Ratten-cDNA (5'-Ende),
O'Hanlon et al.
(1989) J. Biol. Chem. 264: 17389–17394; Wang et al. (1991)
Glycobiology 1: 25–31
- 51. Ratten-Gen (Promotor), Svensson et al. (1990) J. Biol. Chem.
265: 20863–20688
- 52. Ratten-mRNA (Fragmente), Wen et al. (1992) J. Biol. Chem.
267: 2512–2518
-
Zusätzliche
Verfahren und Reagenzien, die in den Verfahren zur Modifikation
der Glycolysierung verwendet werden können, sind in der Literatur
beschrieben, wie das US-Patent Nr. 5,955,422, US-Patent Nr. 4,775,622,
US-Patent Nr. 6,017,743, US-Patent Nr. 4,925,796, US-Patent Nr.
5,766,910, US-Patent Nr. 5,834,251, US-Patent Nr. 5,910,570, US-Patent Nr.
5,849,904, US-Patent Nr. 5,955,347, US-Patent Nr. 5,962,294, US-Patent
Nr. 5,135,854, US-Patent Nr. 4,935,349, US-Patent Nr. 5,707,828
und US-Patent Nr. 5,047,335.
-
Geeignete
Hefe-Expressions-Systeme können
von Quellen wie die American Type Culture Collection, Rockville,
MD erhalten werden. Vektoren sind von einer Vielzahl von Quellen
kommerziell erhältlich.